
Résumés
Abstract
Glacial deposits of the last glaciation associated with the Harvard sublobe (Lake Michigan lobe) in northeastern Illinois, U.S.A., occur between sediment with dateable organics. The lower organics include fragments of Picea sp. as young as 24 000 ± 270 BP. The supraglacial organics occur sparsely in laminated silt and fine sand in landforms that are positioned relatively high on the landscape, such as deposits from ice-walled lakes. These terrestrial organics yield ages that are 2500 to 1300 14C years older than organics at the base of sediment successions in nearby kettle basins. Basal 14C ages from four upland sites range from 17 610 ± 270 to 16 120 ± 80 BP. Our revised time-distance diagram of the Harvard sublobe now reflects a period of stagnation from 24 000 to about 17 600 BP. The supraglacial lacustrine silt yielded plant macrofossil assemblages of primarily tundra plants, including Salix herbacea and Dryas integrifolia. These plants likely grew in supraglacial and ice-marginal environments. The ostracode fauna include Cytherissa lacustris and Limnocythere friabilis. Geomorphic relations and ostracode ecology indicate that more than 17 m of ice buttressed some of the supraglacial lakes.
Résumé
Les sédiments glaciaires associés au sous-lobe de Harvard, dans le Nord-Est de l’Illinois, sont intercalés dans des sédiments dont la matière organique peut être datée. Des fragments de Picea sp. sont présents dans les couches sous-jacentes et remontent au plus à 24 000 ± 270 BP. La matière organique sus-jacente est disséminée dans les limons laminés et les sables fins de dépôts de lacs de barrage glaciaire perchés. Cette matière organique livre des âges 14C qui sont de 2500 à 1300 ans plus vieux que la matière organique de la base des sédiments des marmites glaciaires (kettles). Les âges 14C de quatre sites perchés vont de 17 610 ± 270 à 16 120 ± 80 BP. Notre révision du diagramme distance-temps du sous-lobe de Harvard traduit l’existence d’une période de stagnation glaciaire entre 24 000 et 17 600 BP. Le limon lacustre sus-jacent contient des assemblages macrofossiles de toundra, avec Salix herbacea et Dryas integrifolia. Ces plantes croissaient dans des milieux proches des glaces. Le contexte géomorphologique et l’écologie des ostracodes (Cytherissa lacustris et Limnocythere friabilis) indiquent que certains lacs supraglaciaires étaient étayés par plus de 17 m de glace.
Corps de l’article
Introduction
The advance and retreat of the southern margin of the Laurentide Ice Sheet has been attributed to either climatic forcing factors associated with long-term ENSO cyclicity of about 800 years (Wang et al., 2000, 2003) or to internal glacial mechanisms (Mickelson et al., 1983; Dyke and Prest, 1987; Lowell et al., 1999). Testing hypotheses such as these requires well-dated deposits associated with both glacier advance and retreat. Time-distance diagrams, used to illustrate dynamic ice margins, are often hampered by a few unverified, and in some cases, unreliable radiocarbon ages, especially those obtained from bulk lake sediments (Grimm and Maher, 2002). Inaccuracies will eventually be resolved as key sites are dated by newer techniques such as accelerator mass spectrometry (AMS).
Terrestrial plant and faunal remains are present in deglacial sediments in northeastern Illinois. The aim of this investigation is to describe the sedimentary successions in these settings, describe the flora and fauna, and discuss the significance of the inferred paleoenvironments. The reconstructions indicate that the Harvard sublobe of the Lake Michigan lobe stagnated from about 24 000 to 17 600 BP. Probably the most significant feature of this research is the discovery of terrestrial materials in material not previously known to yield reliable radiocarbon ages, and the potential to more accurately date deglacial successions.
The sites described herein were discovered during mapping of surficial glacial deposits in the urban areas west of Chicago, Illinois (Curry et al., 2001; Grimley and Curry, 2001; Curry, 2005, in review). Most of these sites have been subsequently obliterated by land development and reclamation.
The last glaciation in northeastern Illinois
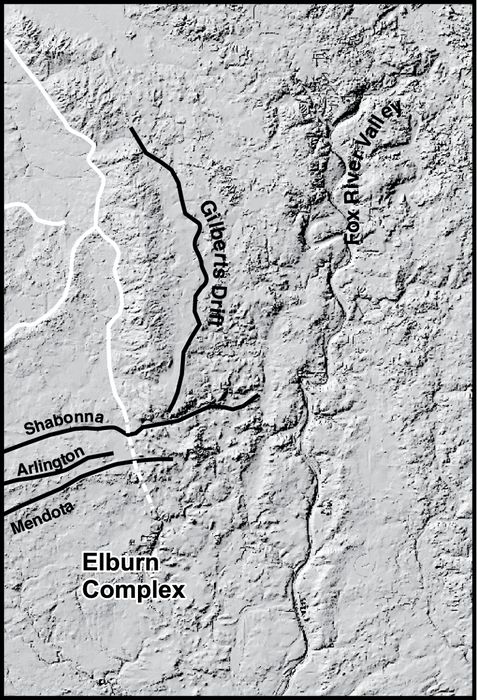
During the last glaciation in northeastern Illinois (the Michigan Subepisode of the Wisconsin Episode), the Lake Michigan lobe was comprised of the Harvard, Princeton, and Joliet sublobes (Fig. 1A; Willman and Frye, 1970). During the Marengo Phase of Hansel and Johnson (1996), the Harvard sublobe formed the Burlington[1] and Marengo Moraines (Fig. 2A). Later, flow of the active portion of the southern Lake Michigan lobe was diverted west by interaction with the Huron-Erie lobe, creating the Princeton sublobe that deposited the Bloomington Morainic System (BMS in Fig. 2A; Wickham et al., 1988). During the Putnam Phase, ice in the Princeton sublobe formed sequentially the Shabonna, Arlington, and Mendota Moraines. Subsequent ice stagnation formed the Elburn Complex (Fig. 2B; Willman and Frye, 1970), an area characterized by kames, kamic hummocks, and eskers. Concurrent stagnation of the Harvard sublobe formed the Gilberts drift (Fig. 2B). During the early part of the Livingston Phase, the Princeton sublobe formed the St. Charles Moraine (Fig. 2C). The southern margin of this landform was influenced by northwesterly flow of the Princeton sublobe (Fig. 1A), but the northern part assumed a north-south trend characteristic of the Joliet sublobe. The difference in moraine orientation likely reflects the melting Huron-Erie lobe. The interaction of these lobes ceased during the late Livingston Phase as suggested by the north-to-south orientation of the Minooka Moraine. The Barlina Moraine was formed by the Harvard sublobe (Fig. 2C) although its continuity with the aforementioned moraines associated with the Livingston Phase was obliterated by fluvial erosion during the subsequent Woodstock Phase.
During the Woodstock Phase, the Harvard and Joliet sublobes formed the Woodstock Moraine and basal portion of the West Chicago Moraine, respectively (Fig. 2D). The Woodstock Moraine is incised by a relatively steep-walled channel eroded by the Fox River, the primary drainage in the eastern half of the study area. Downstream of this breach are several large point bars and an abandoned oxbow-like channel formed by initial erosion of the Woodstock Moraine and subsequent aggradation in the Fox River valley. These features were attributed to the Fox River Torrent by Alden (1904) and Curry (2005) and possibly formed at about the same time as the Kankakee Torrent described by Ekblaw and Athy (1925), Bretz (1940) and Willman and Payne (1942). Deposits that may be associated with the Kankakee/Fox River torrents have been radiocarbon dated at about 16 000 to 15 600 BP (Hajic and Johnson, 1989; Hansel and Johnson, 1996). The last glaciation (Michigan Subepisode) in Illinois ended with the Crown Point Phase (not shown) and formation of the Valparaiso, Tinley, and Lake Border Moraines. Ice finally retreated from Illinois by about 14 000 BP (Hansel and Johnson, 1992).
Sites and methods
Radiocarbon ages of materials collected from our sites and other sites in northeastern Illinois are listed in Table I. Uncalibrated 14C ages are used for discussion purposes; the calibrated values are also given in Table I. Plant macrofossils, ostracodes, and pelecypods were systematically analyzed from monoliths sampled at the Fox River Stone Company (FRSC) and Prairie Pit #94 (PP) sites. Unless otherwise noted, the other sites were cored. Sediment sample volumes of 500 cm3 were sieved for plant macrofossils and insects (nested screens with 425 and 212 µm mesh openings). Pollen samples (1 cm3) were also collected, but poor condition and low concentration of pollen grains resulted in only a representative subsample being analyzed.
Standard procedures were used for the analysis of plant macrofossils (Birks, 1980) and pollen (Faegri and Iverson, 1975). Plant macrofossils were identified with the aid of a collection of seeds, fruits, leaves and other plant organs from modern plants. Species represented in this collection include all those which today inhabit arctic, subarctic, boreal forest, deciduous parkland, and grassland environments. Plant taxonomy and habitat information are based on Welsh (1974), Edlund (1986), Gleason and Conquest (1991), and Aiken et al. (2003).
Samples that were sieved for ostracodes and 14C‑dateable material were pretreated following Forester (1988). Ostracodes were identified by referring to Delorme (1970a, 1970b, 1971). Pelecypod shells were also picked and identified.
Figure 1
A

B

Study area. (A) Location of the three sublobes of the Lake Michigan lobe in northeastern Illinois, and their associated moraines (in gray). (B) Location in context of the major lobes of the Laurentide Ice Sheet in the southwestern Great Lakes region.
Région à l’étude. (A) Carte des trois sous-lobes du lobe du Lac Michigan dans le nord-est de l’Illinois et de leurs moraines respectives (en gris). (B) Carte générale des lobes majeurs de la calotte glaciaire Laurentienne dans la région du sud-ouest des Grands Lacs.
Glacial sediment succession
Older organics
Throughout northeastern Illinois, proglacial lake sediment, outwash, and loess of the last glaciation overlie an organic-rich silt loam to silty loam diamicton known as the Robein Member of the Roxana Silt (Fig. 3). The unit is leached of carbonate minerals and weathered, and is the upper part of the Farmdale Soil, which is typically welded to the Sangamon Soil developed in glacial sediment of the Glasford Formation (Curry, 1989; Curry and Pavich, 1996; Curry et al., 1999). At sites 13, 14, and 15 in northeastern Illinois (Fig. 2A; Table I), tree trunks in growth position and other wood fragments in silt of eolian or lacustrine origin yield ages as young as 23 320 to 24 000 BP (Table I). This age, which marks the end of the Athens Subepisode (a period of primarily loess deposition) and the beginning of the Michigan Subepisode (the last period of glacier activity in northeastern Illinois), is about 1000 14C years younger than what is reported in Hansel and Johnson (1996: Fig. 10). In some places these sediments yield valves of the ostracodes Limnocythere friabilis and Cytherissa lacustris. Fossil plants from these deposits are locally abundant (especially at the Feltes site) (Curry et al., 1999), but have not been identified.
Sediment succession of the last glaciation in northeastern Illinois
Hansel and Johnson (1996) subdivided the glacial succession deposited by the Lake Michigan lobe into two groups. The Wedron Group includes three formations with several members comprised of glacigenic diamicton (typically till) that interfinger with layers of sorted sediment (typically outwash and lake sediment) of the Mason Group, including the aforementioned Roxana Silt (Fig. 3 ; Hansel and Johnson, 1996; Curry et al., 1999).
Younger organics
Supraglacial deposits of fossiliferous, bedded silts and fine sand classified with the Equality Formation occur throughout northeastern Illinois. Here, we describe the stratigraphy and environment of deposition of sediment successions at five main sites with radiocarbon ages (Fig. 4), and provide a brief overview of key additional sites in the region with radiocarbon ages.
Hampshire Quadrangle ice-walled lake deposits
Several core samples from the Hampshire Quadrangle (Curry, in review) indicate that most, if not all, of the circular to subcircular features known as the De Kalb mounds (Flemal et al., 1973) are sediment that were deposited in ice-walled lakes (Iannicelli, 2003; Konen et al., 2005). The genesis of these landforms had been attributed to the disintegration of pingos (Flemal et al., 1973), but new evidence including abundant deep-water ostracodes (described below) and geomorphic associations indicate that some of the deposits were in contact with as much as 17 m of glacial ice (Konen et al., 2005). The loess-mantled ice-walled lake sediment successions include discontinuous layers of sand and gravel that lie between fossiliferous, rhythmically bedded silts and very fine sand (Fig. 4; Curry, in review). Flemal et al. (1973) limited their occurrence in the region to low-lying areas between the Bloomington Morainic System and the Shabonna Moraine (Fig. 2A‑B), but they are probably present on the gently sloping up-ice slopes of most of the moraines.
Figure 2
A
Marengo Phase

B
Putnam Phase
C
Livingston Phase

D
Woodstock Phase

Location of study sites and features noted in text. (A) Landforms associated with the Marengo Phase and sites with pre-glacial organics and the supraglacial ice-walled lake site (H‑3). (B) Landforms associated with the Putnam Phase (black lines) with features identified in Figure 2A (white lines). The Fox River valley is shown, although it was eroded primarily during the Woodstock Phase. (C) Landforms associated with the Livingston Phase (black lines) and older features (white lines). The sites occur on diamicton of the Yorkville Member, Lemont Formation. (D) Landforms associated with the Woodstock Phase (black lines) and older features (white lines). The sites occur on outwash associated with the Haeger Member except for the Randall Road and Sleepy Hollow sites which are associated with aggradation after the Fox River Torrent.
Localisation des sites d’étude et des structures géomorphologiques mentionnées dans le texte. (A) Morphologies associées avec la Phase de Marengo et sites de matières organiques dans les sédiments du lac de barrage glaciaire (H‑3). (B) Morphologies associées avec la phase de Putnam (trait gras) avec les structures identifiées dans la Figure 2A (lignes blanches). La vallée de la rivière Fox est indiquée ; elle a été érodée principalement pendant la Phase de Woodstock. (C) Morphologies associées avec la Phase de Livingston (lignes noires) et les morphologies plus anciennes (lignes blanches). Les sites sont associés au Membre de Yorkville, Formation de Lemont. (D) Morphologies associées avec la Phase de Woodstock et les morphologies plus anciennes (lignes blanches). Les sites sont associés avec le Membre de Haeger sauf pour les sites de Randall Road and Sleepy Hollow qui sont associés avec l’aggradation qui a eu lieu après l’événement torrentiel de la rivière Fox.
Fox River Stone Company site (FRSC)
Exposures along a quarry wall revealed a succession of clayey glacial diamicton and fossiliferous lake sediment mantled by layers of sand and gravel (Fig. 4). The rhythmically bedded lacustrine silt and very fine sand contained abundant plant macrofossils, ostracodes, and fingernail clams. The site was near the crest of the St. Charles Moraine (Fig. 2C) which is formed of the Yorkville Member of the Lemont Formation. The surficial sand and gravel is associated with an outwash fan-delta that was built westward from the Minooka Moraine (Curry et al., 1999).
Prairie Pit #94 site
An exposure in a quarry just northeast of the Fox River Stone Company revealed a succession of surficial, rhythmically bedded silt to medium sand (Fig. 4). The fossiliferous lake sediment is present above silty clay diamicton of the Yorkville Member, and is buried by weathered colluvium and loess. Geomorphic associations between the lake sediment and other features in the quarry suggest that the lake occupied a shallow valley tributary to the Fox River, the main drainage in the region. Bouldery, dolomite-rich outwash deposits associated with a high-level terrace in the valley of the Fox River likely dammed the tributary valley. The lacustrine deposits are therefore slackwater lake deposits. The site is located on the Minooka Moraine.
Table I
Selected radiocarbon ages from northeastern Illinois

N.D. = not determined; N.A. = not applicable
Sites: (1) Mastodon Lake, Phillips Park, Aurora, NW NW Section 35, T. 38 N., R. 8 E.; (2) Brewster Creek, Excavation pit near boring BC‑1, Section 6, T. 40 N., R. 9 E. (Curry et al., in prep.); (3) Crystal Lake, NW Section 6, T. 43 N., R. 8 E.; (4) Sleepy Hollow meander scar, NW NW Section 28 , T. 42 N., R. 8 E.; (5) Bridge boring, Randall Road over Tyler Creek, NW Sect. 9, T. 42 N., R. 8 E.; (6) Nancy Drive, Crystal Lake, SE Section 3, T. 43 N., R. 8 E.; (7) Nelson Lake, NE Section 25, T. 39 N., R. 7 E.; (8) NIU‑123, NE SW Section 22, T. 39 N., R. 9 E. (Springer and Flemal, 1981); (9) Prairie Pit #94, NW Section 26, T. 41 N., R. 8 E.; (10) Fox River Stone Company, NW Section 4, T. 40 N., R. 8 E. (Curry et al., 1999); (11) Sleepy Hollow subdivision flood retention pond, SW Section 6, T. 42 N. R. 8 E.; (12) Ice-walled lake deposit, S 1/2 Section 34, T. 42. N., R. 6 E.; (13) Wedgewood subdivision (temporary exposure at retention pond) NE Section 12, T. 42 N., R. 7 E.; (14) Bluff City mine portal, NE SE Section 30, T. 41 N., R. 9 E.; (15) Feltes sand and gravel pit, NE Section 19, T. 39 N., R. 7 E. (Curry et al., 1999)
Radiocarbon ages <21 000 BP were calibrated using CALIB5 (http://calib.qub.ac.uk/calib/calib.html)
Radiocarbon ages >21 000 BP were calibrated using CALPAL online (http://www.calpal-online.de/); a sigma-one value is given
Sleepy Hollow flood retention pond
Fossiliferous deposits similar to those described at Prairie Pit #94 were exposed during construction of a flood retention pond (Figs. 2C, 4). The primary difference between the sites is the diamicton was that of the Batestown Member (Lemont Formation). The site is located on an eroded portion of the Barlina Moraine (Curry, 2005).
Nancy Drive site
The site is located in a hydrologically closed kettle depression with about 8 m of relief (Figs. 2D, 4). A core from the site includes about 6.4 m of oxidized, fine colluvial sand overlying about 2.3 m of fossiliferous lacustrine silt and silty clay intercalated with thin beds of sandy loam diamicton. The fossiliferous sediment overlies sand and gravel as well as sandy, bouldery diamicton of the Haeger Member. The plant macrofossils from this site were analyzed for 14C dating prior to identification.
Figure 3
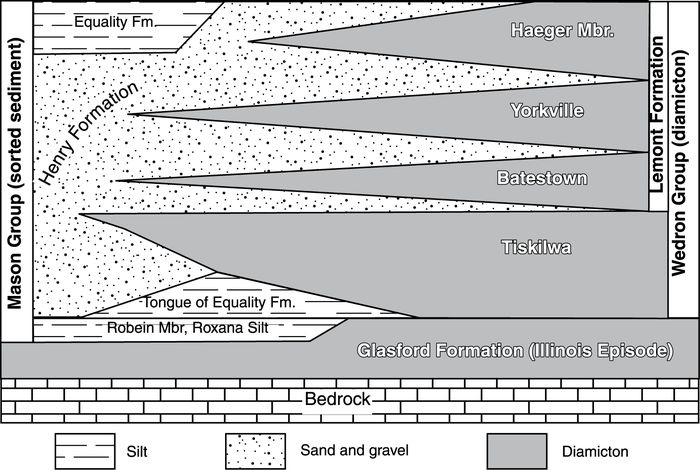
Stratigraphic relations between the Mason and Wedron Groups (after Hansel and Johnson, 1996; Curry et al., 1997b).
Relations stratigraphiques entre le Groupe de Mason et le Groupe de Wedron (d’après Hansel et Johnson, 1996; Curry et al., 1997b).
Randall Road bridge over Tyler Creek
A core from this site revealed a thick succession of fossiliferous slackwater lake sediment (>15 m) in a valley tributary to the Fox River (Fig. 2D). Analysis of the core indicates that the sediment accumulation rate was very high with the two dated intervals separated by approximately 700 years and 9.3 metres of sediment. The fossiliferous lake sediment is present above the Batestown Member.
Sleepy Hollow meander channel
A sediment core from along the channel of a prominent abandoned meander channel revealed thin, fossiliferous lake sediment at a depth of about 1 m, lying above pebbly outwash (Fig. 2D). The radiocarbon ages from this and the Randall Road bridge site (Table I) provide minimum ages for aggradation of outwash in the Fox River valley following the Fox River Torrent.
Nelson Lake and Crystal Lake, Brewster Creek, and Mastodon Lake
Each site occurs in a kettle basin (Fig. 2C‑D). Nelson Lake and Mastodon Lake occur on the flanks of the St. Charles and Minooka moraines, respectively. Crystal Lake and the Brewster Creek site are located on outwash plains associated with the Woodstock Moraine. The radiocarbon ages obtained from fossils in sediments associated with aforementioned ice-marginal environments are from about 1000 to 2750 14C years older than fossils from the kettle sites (Table II).
Fossils
Plant macrofossils
The plant macrofossils from all the sampled sites are well preserved though some leaves have fungal deterioration. A representative selection of these macrofossils is illustrated in Figure 5. In contrast, most of the pollen is extremely degraded (mainly crumpled) and found in low numbers, except for certain samples collected from the PP #94 site where some grains are recognizable and in good condition. The poor pollen recovery precludes quantitative analysis and these data are merely described in the text.
The plant macrofossil assemblages of both the FRSC and PP #94 sites are very similar (Fig. 6) and are considered here as one flora. Plant macrofossils of the primary taxa, described below, were also identified from the sites shown on Fig. 4, with the exception of the Nancy Drive site (where the fossils were not studied).
Arctic tundra plants
The most abundant macrofossils are leaves of Dryas integrifolia (Arctic dryad, white dryad and Arctic avens) (Fig. 5A), Vaccinium uligonosum spp. alpinium (bog bilberry or arctic blueberry) (Fig. 5C), and Salix herbacea (snowbed willow, herb-like willow and dwarf willow). Floral bracts of the latter are shown in Figure 5B. All three species are prostrate dwarf shrubs that form creeping mats. These taxa today inhabit gravelly outwash, wet meadows, and scree slopes in the Arctic (north of 55 °N) and in alpine tundra habitats. The range of Vaccinium uligonosum spp. alpinium extends further south than the other two species into the boreal forest. All three taxa can tolerate both wet and dry soil conditions, but their requirements for soil chemistry differ. Dryas integrifolia and Salix herbacea prefer neutral or alkaline soils, whereas Vaccinium uligonosum spp. alpinium favours acidic soils.
Leaf fragments tentatively identified as those of Rhododendron lapponicum (Lapland rosebay) were only recovered from the FRSC site. This evergreen dwarf shrub has a similar ecology (including soil preference) and geographic range as Vaccinium uligonosum spp. alpinium. Both of these species can not tolerate dense shade, but are found more commonly in open areas or under partial shade in open woodlands.
Figure 4

Lithologic logs of five key sites in northeastern Illinois indicating the relative abundance of ostracodes and plant macrofossils that provided the radiocarbon ages. The corresponding numbers with macrofossil symbols are the uncorrected radiocarbon ages in ka.
Relevés lithologiques de cinq sites clés dans le nord-est de l’Illinois indiquant l’abondance relative d’ostracodes et de plantes fossiles qui fournissent des âges au radiocarbone. Les nombres associés aux symboles de macrofossile sont des âges au radiocarbone non-corrigés en milliers d’années.
Several taxa of arctic herbs were identified from macrofossils. Seeds of a member of Rosaceae (rose family) (Fig. 5D), tentatively identified as Sibbaldia procumbens (sibbaldia) are rare. Like the dwarf shrubs, this ground-hugging perennial herb requires a moist, gravelly soil, and is distributed today across the low (southern) Arctic.
A few calyces of Armeria maritima (sea thrift and sea pink) were also recovered (Fig. 5E). This species is a perennial cushion plant with pink flowers and long, narrow fleshy evergreen leaves (not found as fossils). Armeria maritima exists today in dry sand and gravel, such as found along the coast of oceans and lake shores, and on river terraces. This species is not found under heavy shade.
Seeds of Silene (Fig. 5F), tentatively identified as Silene acaulis (moss campion), are common in the FRSC and PP #94 samples. This is a perennial cushion plant with a woody taproot that forms an extensive moss-like carpet. It colonizes well-drained sandy gravelly and till soils, and inhabits river terraces and shorelines, barrens, and slopes. Silene acaulis has a circumpolar and circumboreal distribution and is found in alpine habitats as far south as New Hampshire.
Also identified were other species which have modern distributions in both the low arctic and boreal forest. An achene of Ranunculus cf. aquatilis (white water-crowfoot and white water-buttercup) (Fig. 5H) was recovered just at the PP #94 site. This annual or perennial plant requires moist or wet soil and can grow in shallow water (<1 m deep), hence it is found today along the edges of streams, ponds and lakes. Although Ranunculus aquatilis is fairly common in Arctic lakes and ponds, it may be occasionally found as far south as the Midwest U.S.A. and New England.
A few drupes of Potamogeton filiformis (slender-leaved pondweed; and filiform pondweed) were found. This species inhabits still or flowing eutrophic fresh water in streams, lakes, and ponds in water depths up to 1 m. It is the only species of pondweed found in the Canadian Arctic Archipelago today (West and Pettit, 2000). Recovered from both sites were florets of a member of Caryophyllaceae (pink/chickweed family), which today inhabits Arctic and boreal habitats.
Table II
Difference in 14C age between ice-marginal sites high in the landscape and basal kettle dates lower in the landscape associated with the same moraine

Subarctic boreal plants
Macrofossils of trees, limited to one species, Picea mariana (black spruce), were recovered in low abundance from the younger (PP #94) site only. One needle of Picea mariana was found (Fig. 5I) as were several stem fragments with woody pegs (for attachment of leaves) identified as Picea sp. (presumably black spruce). This species typically inhabits moist soils, such as bogs, in the boreal forest today.
A species of Brassicaceae (mustard family), possibly that of Cardamine bulbosa (spring cress), was identified (Fig. 5G). This perennial plant requires moist or wet conditions and grows in a range of soil textures. Today Cardamine bulbosa is found in open woods, open wet meadows, and in or near shallow water. It has a modern distribution from the boreal and temperate deciduous forests, as far south as Florida and Texas.
Seeds of Saxifraga sp. (saxifrage) are present in the macrofossil assemblage. These are perennial herbs that grow in a variety of habitats and have primarily a circumboreal distribution. A seed of a boreal forest-type plant, Viola sp. (violet), was also identified.
Pollen
Most of the samples contain few pollen grains that are extremely degraded (particularly crumpled), especially from the FRSC site. Even samples of black laminae from the PP #94 section contained pollen too degraded for quantitative analysis. Maher et al. (1998), Baker et al. (1999) and Heusser et al. (2002) all reported preservation of pollen of full-glacial age in some sediment samples, but not in others, even though they were all collected from the same monoliths. Explanations for poor pollen recovery in these sites include: pollen concentrations being diluted during times of rapid loess deposition and other glacigenic sediment (e.g. Heusser et al., 2002); tundra plants being poor pollen producers, since many are insect-pollinated (Birks, 1980; Gajewski et al., 1995); and oxidization of pollen grains within sediment pores (Cushing, 1967; Hall, 1981).
Pollen grains of a few taxa, those of Picea glauca-type (white spruce), Abies (fir), and Ambrosia-type (ragweed), are moderately well preserved. But only those of spruce are common, suggesting that spruce, probably Picea glauca, was present somewhere in the region during this time. Considering the low pollen counts of this taxa per microscope slide, it is doubtful that white spruce existed in the local area. Abies probably also existed in the region (though not in the immediate area), since experimental studies of modern pollen deterioration in soils indicate that grains of this taxon deteriorate readily after deposition (Hall, 1981).
Ostracodes
Ostracodes were recovered from most sites described above except for the Nancy Drive and PP #94 sites. Their absence from these sites may be attributed to the low total dissolved solid content of the host water in tandem with slow sediment accumulation rates. Alternatively, methanogenesis may be invoked as a cause for shell dissolution (Jones et al., 1982; Curry et al., 1997a).
Limnocythere friablis (Fig. 7A, D) was the most abundant ostracode identified in the ice-marginal and supraglacial lake sediment. This species is among the smallest in North America (Delorme, 1971) with the average valve length reaching about 55 µm. Only females were observed, suggesting that in these extreme ice-marginal environments the species reproduced via parthenogenesis. Only three of more than 5 500 modern freshwater aquatic sites in Canada yielded Limnocythere friabilis (Denis Delorme, personal communication). The species is also present in Lake Michigan at water depths between 30 and 75 m (Buckley, 1975), and in Lake Erie at depths below about 15 m (Benson and MacDonald (1963). Limnocythere friabilis is also abundant in Pleistocene lake sediments of Lake Michigan (Staplin, 1963; Forester et al., 1994) and early postglacial kettle successions in Illinois such as the one described at Nelson Lake (Curry, 2003).
Cytherissa lacustris (Fig. 7B, E) was also abundant, particularly in the ice-walled lake deposits. This species prefers living in relatively dilute water ranging in total dissolved solids from about 10 to 250 mg/L (Delorme, 1978, 1989). Cytherissa lacustris is found commonly throughout Canada in waters greater than 3 m depth surrounded by boreal vegetation. Its upper temperature tolerance is about 23 °C. Like Limnocythere friabilis, Cytherissa lacustris is found in the profundal zone of the modern Great Lakes at water depths ranging from 15 to 45 m (Benson and MacDonald, 1963; Buckley, 1975; Delorme, 1978).
Figure 5
A
Dryas integrifolia

B
Salix herbacea

C
Vaccinium uligonosum spp. alpinium

D
Sibbaldia procumbens

E
Armeria maritima

F
Silene cf. S. acaulis

G
cf. Cardamine bulbosa

H
Ranunculus cf. R. aquatilis

I
Picea mariana

Scanning Electron Microscope (SEM) micrographs of selected plant macrofossils. Note the reference scales (white line) are of various lengths.
Sélection de photographies des plantes macrofossiles obtenues à partir d’un microscope électronique à balayage. Notez que les échelles de référence sont de longueur variable.
Figure 6
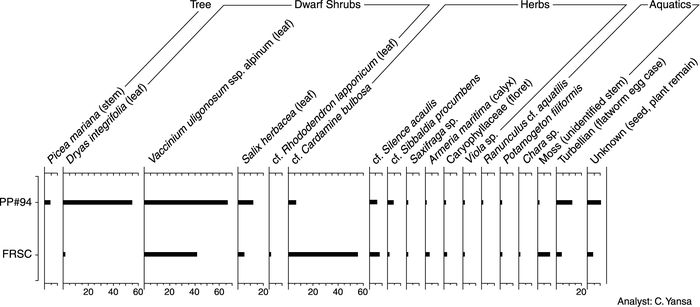
Plant macrofossil abundance diagram showing counts for both the Fox River Stone Company (FRSC) (older) and Prairie Pit #94 (PP #94) (younger) sites.
Diagramme de comptage des plantes macrofossiles pour le site Fox River Stone Company (FRSC) (plus ancien) et le site Prairie Pit #94 (PP #94) (plus jeune).
Limnocythere herricki (Fig. 7C, F, I) is known from the prairies or within the prairie-forest transition in Canada where long cold winters (205 to 224 frost days), warm to cool summers, and high drought frequency are common (Forester et al., 1987a).
Candona rectangulata (Fig. 7H) is present in freshwater lakes located in the northern boreal forest and tundra of northeastern Canada north of 65 °N latitude. This species is tolerant of high salinity, but it cannot tolerate sustained temperatures above 15 °C (Forester et al., 1987b).
Paleohydrological interpretation
The ostracode assemblages of the ice-marginal sites discussed in this paper are dominated by Limnocythere friabilis and Cytherissa lacustris, with fewer Limnocythere herricki, Candona rectangulata, and Heterocypris incongruens. At the FRS site, Limnocythere friabilis dominates in the lower part of the succession, whereas the Pisdium sp. clams are more abundant in the upper part of the record along with the other ostracode species, notably Limnocythere herricki (Fig. 8). In the ice-walled lake deposits, Cytherissa lacustris and Limnocythere friabilis are most abundant. Specific interpretations are difficult due to lack of published data on ostracode occurrences in specific types of arctic and sub-arctic lakes. The two most common ostracodes co-occur in the southern Great Lakes at water depths approaching or below the thermocline between 15 to 45 m. The epilimnion of the ancient, smaller, ice-marginal lakes in northeastern Illinois probably warmed rapidly in the summer, but were likely heated less intensely and for a shorter time than the Great Lakes are today. Cytherissa lacustris indicates temperatures may have been as great as 23 °C. The observation that the most abundant fossil ostracode (Limnocythere friabilis) is only abundant in relatively shallow zones of large lakes (Forester et al., 1994) suggests that this species thrived in lakes that discharged meltwater or (cold) groundwater.
Other invertebrates
Arthropod remains are uncommon and limited to two types. Thoracic segments of Caddisfly (Order Trichoptera) larvae were recovered. These larvae are known to live in cases on the underside of rocks in streams and along lake shores (Borror et al., 1989). Also recovered were cocoons of Turbellaria (flatworm), which today inhabit laminated lake sediment (Borror et al., 1989). Shells of Pisidium sp. (fingernail clams) also were found in certain stratigraphic positions and indicate sediment was deposited in relatively quiet water.
Discussion
Paleoenvironments of northeastern Illinois
In the ice-walled lake environments studied in Illinois, plants grew on sandy and gravelly soils that developed directly on the glacial ice and were washed or collapsed into the lakes as the underlying ice melted. The slackwater lake and proglacial lake environments represented at the FRSC and PP #94 sites, respectively, contain many ostracodes and plant fossils common to the ice-walled lakes. Other environments, such as relatively ice-free outwash deposits or silty abandoned strand deposits, were likely available for colonizing by plants. Only a few taxa, such as Potamogeton filiformis (slender-leaved pondweed) and possibly Ranunculus cf. aquatilis (white water-crowfoot), occupied the shallow water along the lakes. The absence of an aquatic-emergent flora and the limited suite of submerged aquatic plants suggest that the lakes were oligotrophic.
Figure 7
A
Limnocythere friabilis, female, left valve

B
Cytherissa lacustris, left valve

C
Limnocythere herricki, male, left valve
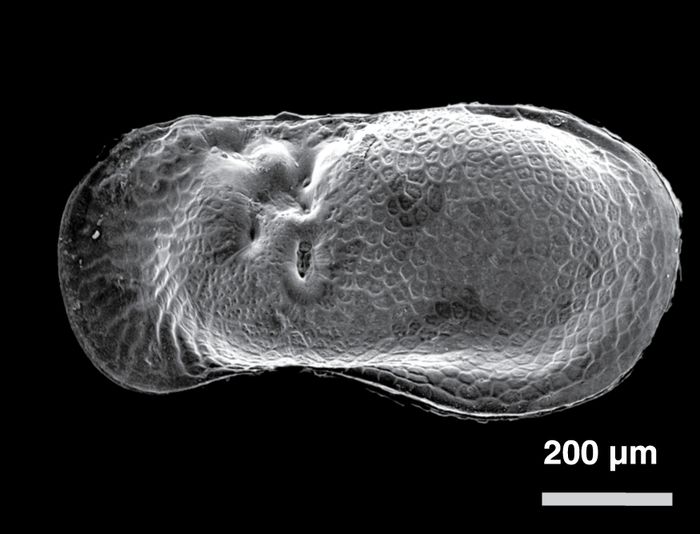
D
Limnocythere friabilis, female, right valve

E
Cytherissa lacustris, right valve

F
Limnocythere herricki, male, right valve

G
Heterocypris incongruens, female, left valve

H
Candona subtriangulata, female, right valve

I
Limnocythere herricki, male, right valve
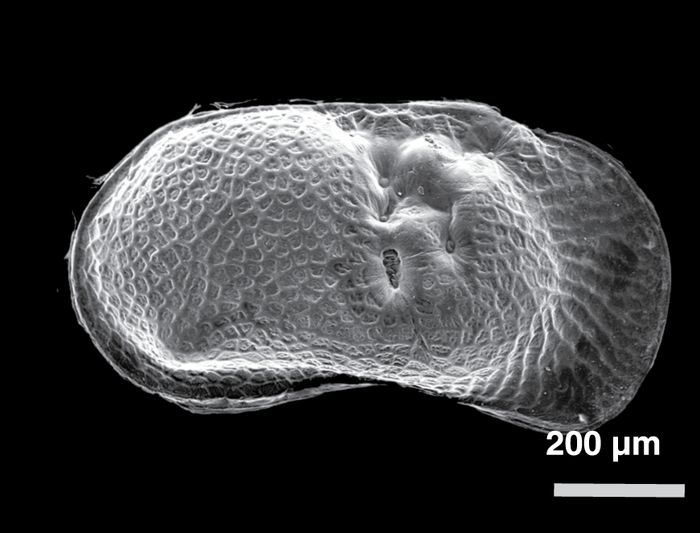
Scanning Electron Microscope (SEM) micrographs of selected ostracodes.
Sélection de photographies d’ostracodes obtenues à partir d’un microscope électronique à balayage.
Figure 8

Relative abundance of ostracode valves and Pisidium clam shells from the Fox River Stone Company monolith.
Abondance relative des valves d’ostracode et de Pisidium du monolithe de Fox River Stone Company.
The abundance of Vaccinium uligonosium spp. alpinium and Salix herbacea (snowbed willow) leaves in the region may not just be attributed to the commonness of these dwarf shrubs, but also because they are deciduous. Leaves of Dryas integrifolia (Arctic dryad), in contrast, are evergreens and hence may be under-represented in the fossil record. These and most other species identified are found in recent fluvial deposits of the Canadian Arctic (West and Pettit, 2000). Thus, the deposition of modern plant remains is not spatially uniform and some species are over‑ or under-represented in the fossil record relative to their actual abundance (West and Pettit, 2000).
Ecological studies also reveal that both Salix herbacea and Dryas integrifolia are pioneers on stony and gravelly habitats, such as recently exposed river flats and lake shores, where they stabilize the substrate with long taproots and fix nitrogen via the ectomycorrhizal nodules attached to their roots. Dryas integrifolia can not endure competition from other plants for space, so its abundance at the study sites further substantiates our interpretation of a tundra-like environment. Dryas integrifolia is also commonly found today with Silene acaulis (moss campion), which is a perennial cushion plant that has been tentatively identified at our sites. Both of these taxa are reported in ecological studies to withstand open-tundra and drying alpine winds on mountain tops. Sibbaldia procumbens (sibbaldia) is another pioneer tentatively identified in our study. Today, this perennial herb is commonly found with Salix herbecea under snow patches that persist late into the Arctic summer.
We interpret the paleovegetation as “tundra-like” rather than “tundra”, because of the presence in the fossil assemblage of a few taxa which today have ranges south of the tree-line. Species which are found in the boreal forest and the Arctic today include Silene acaulis, Ranunculus cf. aquatilis (white water-crowfoot), and Potamogeton filiformis (slender-leaved pondweed) (Porsild and Cody, 1980). The remainder of the flora, Picea mariana (black spruce), cf. Cardamine bulbosa (spring cress), Saxifraga sp. (saxifrage), and Viola sp. (violet), are presently restricted to the boreal forest zone.
The rare occurrence of Picea mariana macrofossils, and absence of pollen of this taxon suggest that black spruce was probably a minor constituent of this flora. Moreover, these fossils were only recovered from the younger PP #94 site, suggesting that black spruce arrived in the study area after 16 500 BP. In addition to climate, the organic-poor soils, as indicated by the shrub and herb taxa, would also have limited the range of Picea mariana (black spruce) which requires richer soils. Also, Birks (1976) suggested that strong winds during the full-glacial may have been more effective than low temperatures in limiting tree growth. Black spruce probably grew on moist soils once summer temperatures warmed above the July 10 °C isotherm that today delineates tree-line (Ritchie and Harrison, 1993), and when high winds diminished. Even though well-preserved pollen grains of Picea glauca-type (white spruce) were recovered, the biased pollen assemblages, low pollen concentrations, and the absence of Picea glauca macrofossils suggest that white spruce was not present in the immediate area.
Comparison with other coeval fossil localities
The ages assigned to many sites with tundra-like vegetation are probably inaccurate because the 14C chronologies are based on the dating of bulk organic sediment (gyttja and marl) (e.g. Baker, 1965; Birks, 1976). These sediments are now known to provide ages that are erroneously older by at least a millennium, because of the “hard-water effect” (MacDonald et al., 1991). For example, pollen and plant macrofossils from the Wolf Creek site in central Minnesota were interpreted as indicative of a full-glacial tundra environment from 20 000 to 14 000 BP (Birks, 1976). While the interpretations regarding paleovegetation are reasonable, the chronology is too old and predates the now accepted deglaciation chronology for this part of Minnesota. Our 14C chronology, based on terrestrial plant macrofossils, provides a more accurate determination of the timing of deglacial events and vegetation changes. With reference to other studies, we reject all ages derived from sediment and other aquatic materials and only report those obtained from terrestrial plant macrofossils.
Most researchers who interpret interstadial and late-glacial vegetation from pollen stress the importance of long-traveled pollen in the records. McAndrews (1984) reported current deposition of deciduous tree pollen on a glacier in the Canadian Arctic and determined that these grains had traveled over 1 000 km. Birks (2003) stressed the importance of studying plant macrofossils when reconstructing interstadial and late-glacial vegetation and climate because of the problems of long-distance dispersal of pollen and over-representation of tree pollen in Arctic environments. Our results support this conclusion since we recovered Picea mariana needles of local origin, but pollen grains, tentatively identified as those of Picea glauca, were likely far-traveled. Similarly, pollen grains of Picea (spruce) and deciduous trees were found associated with “Arctic” macrofossils, such as Salix herbacea and Dryas integrifolia, at both Spider Creek (Baker, 1965), and Wolf Creek (Birks, 1976) in Minnesota, and at the Valders Quarry in Wisconsin (Maher et al., 1998). Likewise, they interpreted that the pollen grains were derived from distal sources.
Figure 9
A

B

Time-distance diagrams for the active margin of the Harvard sublobe, northeastern Illinois. See Figure 1A for the line of section. (A) Diagram based on the radiocarbon ages in Springer and Flemal (1981), Curry et al. (1997a), Curry et al. (1999). (B) Revised time-distance diagram based on the new radiocarbon ages (Table I). We reject the ages from the Wedgewood and NIU‑123 sites (see discussion).
Diagramme de distance-temps pour la marge active du sous-lobe de Harvard, nord-est de l’Illinois. Se référer à la Figure 1A pour la localisation de la section. (A) Diagramme basé sur les âges au radiocarbone dans Springer et Flemal (1981), Curry et al. (1997a), Curry et al. (1999). (B) Diagramme de distance-temps révisé basé sur les nouveaux âges au radiocarbone (Table I). Nous excluons les âges provenant des sites de Wedgewood et de NIU‑123 (voir la discussion).
The nearest site comparable in age to ours is that of the Conklin Quarry in southeastern Iowa, which dates from 18 000 to 16 700 BP (Baker et al., 1986). The site is located 250 km west of the study area. The pollen assemblage is dominated by Picea (spruce), Pinus (pine), and Cyperaceae (sedge family) pollen and associated with macrofossils of spruce with those of dwarf shrubs and tundra herbs (Baker et al., 1986). The Conklin Quarry site macrofossil taxa are similar to our sites, including Dryas integrifolia and Picea mariana, but lacking Salix herbacea. From the plant and insect fossils they estimated that the mean summer temperature was 11‑13 °C cooler than at present. Farther south, macrofossils and pollen records suggest that a broad-leaved/evergreen mixed forest was present in the lower Mississippi Valley at 35 °N at about 15 000 BP (Jackson et al., 2000).
In Minnesota, the macrofossil assemblages from the base of cores from Spider Creek (Baker, 1965) and Wolf Creek (Birks, 1976) are nearly identical to what we identified in this study. Our sites, however, lack macrofossils of Carex (sedge), Stellaria (chickweed), Arenaria (sandwort), Juncus (rush), Larix laricina (tamarack), and Poaceae (grass). A site in southeastern Minnesota revealed that Arctic (e.g. Dryas integrifolia and Salix cf. herbacea) and subarctic vegetation existed on a treeless or nearly treeless landscape at about 18 700 BP (Baker et al., 1999). The pollen spectrum of this site included Picea (spruce), Pinus (pine), and Cyperaceae (sedge family), which was interpreted as allochthonous.
To the north of our study area, similar open-tundra vegetation was established in eastern Wisconsin (Valders Quarry) after the retreat of the Green Bay lobe at about 14 500 BP (Maher et al., 1998). Interestingly, the ostracode fauna are also nearly identical, with only the addition of the deep-water cryophyllic ostracode Candona subtriangulata. This is the only site from the southern Great Lakes region with a well-described interstadial ostracode fauna. At the Two Creeks site in northeastern Wisconsin, Kaiser (1994) indicates that the Two Creekan Interstade persisted from 12 050 to 11 750 BP. During this time Picea mariana (black spruce) occupied the swampy soils along the shore of glacial Lake Chicago when the lake was at or just below the present level of Lake Michigan (Kaiser, 1994). These ages are comparable to the date of 12 050 BP obtained from Salix sp. (willow) stems, associated with several mosses and leaves of Dryas integrifolia and Salix herbacea, at the Cheboygan Bryophyte Bed in northern Lower Michigan (Larson et al., 1994).
Tundra localities outside the upper Midwest
Comparison of tundra-like fossil localities in eastern North America with those from the upper Midwest provide valuable insight into deglacial environmental conditions along the southern margin of the Laurentide Ice Sheet. No tundra-like fossil sites dating to the full‑ or late-glacial have yet been found on the northern Great Plains west of Wolf Creek, Minnesota. Yansa (2006) attributed this to the late deglaciation of this region; by the time active ice had retreated, white spruce trees were already present in the immediate area.
Tundra fossils have been reported at several sites in Ontario (e.g. Terasmae et al., 1966; Morgan and Morgan, 1979; Anderson, 1982; McAndrews and Jackson, 1988; Warner et al., 1988; Karrow, 1990), but most of these have poor chronological control. One exception is Yu (2003) recent study of macrofossil assemblages from the base of cores from Crawford Lake and Twiss Marl Pond in southern Ontario. He reported initial colonization by sparse tundra vegetation comprised of Alnus (alder), Dryas integrifolia, Salix herbacea, and Cyperaceae (sedge family) with some boreal plants at an estimated age of 13 000 BP (extrapolated down core from a date of 10 920 BP). By about 12 000 BP, the vegetation was dominated by Picea, suggesting that the oldest parts of the macrofossil records at these sites represent a transition from tundra (or tundra-like) vegetation to boreal forest.
Macrofossil analysis indicates that Dryas integrifolia, Salix sp. (willow) and Carex sp. (sedge) inhabited south-central Pennsylvania by 14 400 BP where they are assumed to be coeval with local periglacial features (Delano et al., 2002). In Quebec, New York, and New England there are scattered fossil localities bearing full-glacial and late-glacial assemblages of tundra plants with a minor component of boreal vegetation, such as Picea and/or Populus balsamifera (balsam poplar) (e.g. Argus and Davis, 1962; Miller and Thompson, 1979; Richard, 1979; Davis and Jacobson, 1985; Miller and Spear, 1999). Only about half of the taxa that we identified at our sites were also found at these eastern sites. This suggests that during the full-glacial and late-glacial a fairly uniform tundra-like flora existed within the upper Midwest and that it shared some characteristics with a coeval flora that existed in eastern North America. Dryas integrifolia, of note, is found at all of these sites, which agrees with Porsild (1947) that this species was common and widespread in postglacial time.
Revised time-distance diagram for the Harvard sublobe
Curry et al. (1997a) used a radiocarbon age from the Wedgewood site of 23 230 ± 550 BP (Fig. 2A; Table I) to document the interstadial Wedgewood Phase (Fig. 9A). The age was obtained on bulk, fine organics washed from a lacustrine deposit that was present above diamicton of the Tiskilwa Formation, and below outwash associated with the Haeger Member. The deposit also contained abundant Cytherissa lacustris and Limnocythere friabilis. With the new AMS dates obtained from plant macrofossils from the ice-walled lake deposits, it seems likely that the age from the Wedgewood site is contaminated with old carbon.
The new radiocarbon ages suggest that the Harvard sublobe stagnated for about 4000 14C years. The onset of stagnation began at an unknown point (about 21 000 BP) when ice activity in Harvard sublobe stopped after formation of the Marengo Moraine (Fig. 9B). The Harvard sublobe was starved for new ice at this time because the direction of ice flow was diverted by interaction of the Lake Michigan lobe with the Huron-Erie lobe (Wickham et al., 1988). Ice activity in the Harvard sublobe resumed at about 18 000 BP when the St. Charles and possibly Barlina moraines began to form. The similarity in the basal ages of four sites with tundra plant macrofossils indicate that the Harvard sublobe melted fairly uniformly across an area comprising the Burlington Moraine, Marengo Moraine, Gilberts Drift, the eastern parts of the Shabonna, Arlington, and Mendota moraines, as well as the Elburn Complex and St. Charles Moraine. The age at the Fox River Stone Company of 17 540 ± 130 (Fig. 4) represents when the Harvard sublobe reactivated and the initial activity of the Joliet sublobe. The orientation of the southern part of the St. Charles Moraine (Fig. 2C) suggests its formation was influenced by interaction with the Huron-Erie lobe. By contrast, the orientation of the younger Minooka Moraine indicates that interaction between the Lake Michigan and Huron-Erie lobes had ceased.
The age of the large glaciofluvial landforms in the Fox River valley, and hence, the age of the Fox River Torrent during the Milwaukee Phase, is limited by the age from the Nancy Drive site on the Woodstock Moraine at about 14 900 BP and the ages from the Sleepy Hollow subdivision and Prairie Pit #94 sites at about 15 700 BP. The Fox River Torrent may have occurred at about the same time as the Kankakee Torrent (Willman and Payne, 1942).
Conclusions
From about 21 000 to 18 000 BP, the Harvard sublobe stagnated in northeastern Illinois. In the latter stages of melting, slackwater and ice-walled lake environments preserved fossils of tundra vegetation and lacustrine organisms. Such environments are ubiquitous in continentally-glaciated areas (Menzies and Shilts, 1996). For example, there are more than 500 ice-walled lake deposits in northeastern Illinois (the De Kalb Mounds of Flemal et al., 1973), and each one examined so far has yielded plant macrofossils. Younger fossiliferous ice-walled lake deposits dating from the Younger Dryas occur in North Dakota (Clayton, 1967; Tuthill et al., 1964). We hope that these and the previous results encourage others to examine deglacial landforms in other areas not only for paleoecological reconstructions, but for their potential for unraveling deglacial histories.
The oldest ages marking the onset of deglaciation in northeastern Illinois are from terrestrial fossils preserved in the sediment of ice-walled lakes and slackwater lakes. These deposits often occur in areas relatively high on the landscape. Prior to this study, the oldest reliable deglacial ages came from fossils preserved in the base of kettle successions. We have found that such ages are from 1000 to 2500 14C years younger than dated material from ice-walled lakes and slackwater lake deposits located on the same moraines or their outwash plains (Table II). The difference is attributed to the relatively rapid downwasting of surficial ice that formed the ice-walled and slackwater lakes compared to the delayed and slower formation of kettles which involved the melting of buried ice.
From 17 500 to 15 700 BP, the supraglacial and ice-marginal terrestrial environments in northeastern Illinois were dry and windy, as suggested by plant macrofossils of dwarf tundra shrubs and low Arctic herbs, such as Salix herbacea, Dryas integrifolia, Vaccinium uligonosum spp. alpinium, and Silene cf. acaulis. A few plants with boreal forest affinities also were identified, resulting in our interpretation of an anomalous “tundra-like” environment, rather than “tundra”. Many, if not all sites were devoid of trees judging from the general absence of needles and twigs, and the presence of tundra plants that cannot live under a forest canopy such as Armeria maritima. Deposits in northeastern Illinois dating from about 17 500 to 14 200 BP do not contain wood fragments larger than a few millimetres in diameter that are representative of the deposit’s age; radiocarbon ages of larger wood fragments from such deposits invariably yield dates >24 000 BP, indicating reworking of fossils from the Robein Silt.
Parties annexes
Acknowledgements
We thank the following individuals for their identifications: Richard G. Baker, University of Iowa (plant macrofossils); Barry Miller, Kent State University (fingernail clams); Alan Ashworth, North Dakota State (insects) and Patricia Sanford, University of Wisconsin-Madison (flatworm cocoons). David Grimley, Illinois State Geological Survey, assisted in sample collection from the FRS site. Eric Grimm, Illinois State Museum, helped core several of the sites. André Pugin did the French translations. Publication was authorized by the Chief, Illinois State Geological Survey. We thank the reviewers for their helpful comments, especially Andrew Stumpf and Ardith Hansel.
Note
-
[1]
The Burlington moraine is the informal name given to an arcuate ridge in northeastern De Kalb and northwestern Kane County. The moraine is formed of the Tiskilwa Formation, and was previously mapped as part of the Elburn Complex by Willman and Frye (1970). The village of Burlington is located near the centre of the moraine.
References
- Aiken, S.G., Dallwitz, M.J., Consaul, L.L., McJannet, C.L., Gillespie, L.J., Boles, R.L., Argus, G.W., Gillett, J.M., Scott, P.J., Elven, R., LeBlanc, M.C., Brysting, A.K. and Solstad, H., 2003. Flora of the Canadian Arctic Archipelago: Descriptions, Illustrations, Identification, and Information Retrieval. Version: 29 April 2003. http://www.mun.ca/biology/delta/arcticf/.
- Alden, W.C., 1904. The Delavan Lobe of the Lake Michigan glacier of the Wisconsin state of glaciation and associated phenomena. U.S. Geological Survey, Professional Paper 34, 106 p.
- Anderson, T.W., 1982. Pollen and plant macrofossil analyses on late Quaternary sediments at Kitchener, Ontario. Geological Survey of Canada, Ottawa, Current Research 82‑1A : 131‑136.
- Argus, G.W. and Davis, M.B., 1962. Macrofossils from a late-glacial deposit at Cambridge, Massuchusetts. American Midland Naturalist, 67 : 106‑117.
- Baker, R.G., 1965. Late-glacial pollen and plant macrofossils from Spider Creek, southern St. Louis County, Minnesota. Geological Society of America Bulletin, 76 : 601‑610.
- Baker, R.G., Mason, J.A. and Maher, L.J., 1999. Petaloid organs preserved in an arctic plant macrofossil assemblage from full-glacial sediments in southeastern Minnesota. Quaternary Research, 52 : 388‑392.
- Baker, R.G., Rhodes II, R.S., Schwert, D.P., Ashworth, A.C., Prest, T.J., Hallberg, G.R. and Janssens, J.A., 1986. A full-glacial biota from southeastern Iowa, USA. Journal of Quaternary Science, 1 : 91‑107.
- Benson, R.H. and MacDonald, H.C., 1963. Postglacial (Holocene) ostracodes from Lake Erie. University of Kansas Paleontological Contributions, Article 4, 26 p.
- Birks, H.H., 1980. Plant macrofossils in Quaternary lake sediments. Ergebnisse der Limnologie, Archiv für Hydrobiologie 15, 60 p.
- Birks, H.H., 2003. The importance of plant macrofossils in the reconstruction of late glacial vegetation and climate: examples from Scotland, western Norway, and Minnesota, USA. Quaternary Science Reviews, 22 : 453‑473.
- Birks, H.J.B., 1976. Late-Wisconsinan vegetational history at Wolf Creek, central Minnesota. Ecological Monographs, 46 : 395‑429.
- Borror, D.J., Triplehorn, C.A., and Johnson, N.F., 1989. An Introduction to the Study of Insects, 6th edition. Saunders College Publishing, Philadelphia, 875 p.
- Bretz, J.H., 1940. Geology of the Chicago region. Illinois State Geological Survey, Bulletin 65, Part I, 53 p.
- Buckley, S.B., 1975. Study of post-Pleistocene ostracode distribution in the soft sediments of southern Lake Michigan. Ph.D. thesis, University of Illinois, Urbana-Champaign, 293 p.
- Clayton, L., 1967. Stagnant-glacier features of the Missouri Coteau in North Dakota, p. 25‑46. In L. Clayton and T.F. Freers, eds., Glacial Geology of the Missouri Coteau and Adjacent Areas, North Dakota Geological Survey, Miscellaneous Series 30, 170 p.
- Curry, B.B., 1989. Absence of Altonian glaciation in Illinois. Quaternary Research, 31 :1‑13.
- Curry, B.B., 2003. Linking Ostracodes to Climate and Landscape, p. 223‑246. In L.E. Park and A.J. Smith, eds., Bridging the Gap: Trends in the Ostracode Biological and Geological Sciences, The Paleontological Society Papers 9, 290 p.
- Curry, B.B., 2005. Surficial geology of Crystal Lake Quadrangle, McHenry and Kane Counties, Illinois. Illinois State Geological Survey, Illinois Geological Quadrangle Map, IGQ‑Crystal Lake‑SG, Scale 1:24 000.
- Curry, B.B., In review. Surficial geology of Hampshire Quadrangle, Kane and De Kalb Counties, Illinois, Illinois State Geological Survey, Illinois Geological Quadrangle Map, IGQ‑Hampshire‑SG, Scale 1:24 000.
- Curry, B.B. and Pavich, M.J., 1996. Absence of glaciation in Illinois during marine isotope stages 3 through 5. Quaternary Research, 31 : 19‑26.
- Curry, B.B., Anderson, T.F. and Lohmann, K.C., 1997a. Unusual carbon and oxygen isotopic ratios of ostracodal calcite from last interglacial (Sangamon Episode) lacustrine sediment in Raymond Basin, Illinois, USA. Journal of Paleolimnology, 17 : 421‑435.
- Curry, B.B., Berg, R.C. and Vaiden, R.C., 1997b. Geologic mapping for environmental planning, McHenry County, Illinois. Illinois State Geological Survey, Circular 559, 79 p.
- Curry, B.B., Grimley, D.A., and Stravers, J.A., 1999. Quaternary geology, geomorphology, and climatic history of Kane County, Illinois. Illinois State Geological Survey, Guidebook 28, 40 p.
- Curry, B.B., D.A. Grimley, and T.H. Larson. 2001. Surficial geology of Sugar Grove Quadrangle, Kane County, Illinois: Illinois State Geological Survey, Illinois Geological Quadrangle Map, IGQ‑Sugar Grove‑SG. 1:24 000
- Cushing, E.J., 1967. Evidence for differential pollen preservation in Late Quaternary sediments in Minnesota. Review of Palaeobotany and Palynology, 4 : 87‑101.
- Davis, R.B. and Jacobson Jr., G.L., 1985. Late glacial and early Holocene landscapes in northern New England and adjacent areas in Canada. Quaternary Research, 23 : 341‑368.
- Delano, H.L., Miller, N.G. and Potter Jr., N., 2002. Plant fossil evidence for late Pleistocene tundra conditions in south-central Pennsylvania. Abstract with Programs, Geological Society of America, 34 : A‑27.
- Delorme, L.D., 1970a. Freshwater ostracodes of Canada. Part II. Subfamily Cypridopsinae and Herpetocypridinae, and family Cyclocyprididae. Canadian Journal of Zoology, 48 : 253‑266.
- Delorme, L.D., 1970b. Freshwater ostracodes of Canada. Part III. Subfamily Candonidae. Canadian Journal of Zoology, 48 : 1099‑1127.
- Delorme, L.D., 1971. Freshwater ostracodes of Canada. Part V. Families Limnocytheridae, Loxoconchidae. Canadian Journal of Zoology, 49 : 43‑64.
- Delorme, L.D., 1978. Distribution of freshwater ostracodes in Lake Erie. Journal of Great Lakes Research, 4 : 216‑220.
- Delorme, L.D., 1989. Methods in Quaternary Ecology #7. Geoscience Canada, 16 : 85‑90.
- Dyke, A.S. and Prest, V.K., 1987. Late Wisconsinan and Holocene history of the Laurentide ice sheet. Géographie physique et Quaternaire, 41 : 237‑263.
- Edlund, S., 1986. Common Arctic Wildflowers of the Northwest Territories. Department of Indian and Northern Affairs, Yellowknife, Northwest Territories, 32 p.
- Ekblaw, G.E. and Athy, L.F., 1925. Glacial Kankakee torrent in northeastern Illinois. Geological Society of America Bulletin, 26 : 417‑427.
- Faegri, K. and Iverson, J., 1975. Textbook of Pollen Analysis. Hafner Press, New York. 295 p.
- Flemal, R.C., Hinkley, K.C. and Hesler, J.L., 1973. De Kalb Mounds: A possible Pleistocene (Woodfordian) pingo field in north-central Illinois, p. 229‑250. In R.F. Black, R.P. Goldthwait, and H.B. Willman, eds., 24 000, The Wisconsinan Stage. Geological Society of America, Memoir 136, 334 p.
- Forester, R.M., 1988. Nonmarine calcareous microfossil sample preparation and data aquisition procedures. United States Geological Survey, Technical Procedure HP‑78 RI, p. 1‑9.
- Forester, R.M., Delorme, L.D. and Bradbury, J.P., 1987a. Mid-Holocene climate in northern Minnesota. Quaternary Research, 28 : 263‑273.
- Forester, R.M., Delorme, L.D., and Ager, T.A., 1987b. A lacustrine record of late Holocene climate changes from south-central Alaska. Geophysical Monographs, 55 : 33‑40.
- Forester, R.M., Colman, S.M., Reynolds, R.L. and Keigwin, L.D., 1994. Lake Michigan’s Late Quatenary limnological and climate history from ostracode, oxygen isotopes and magnetic susceptibility. Journal of Great Lake Research, 20 : 93‑107.
- Gajewski, K., Garneau, M. and Bourgeois, J.C., 1995. Paleoenvironments of the Canadian High Arctic derived from pollen and plant macrofossils: problems and potentials. Quaternary Science Reviews, 14 : 609‑629.
- Gleason, H.A. and Conquest, A., 1991. Manual of Vascular Plants of Northeastern United States and Adjacent Canada, 2nd edition. New York Botanical Garden, Bronx, New York, 910 p.
- Grimley, D.A. and Curry, B.B., 2001. Surficial geology of Geneva Quadrangle, Kane County, Illinois. Illinois State Geological Survey, Illinois Geological Quadrangle Map, IGQ‑Geneva‑SG, Scale 1:24 000.
- Grimm, E.C. and Jacobson Jr., G.L., 2004. Late-Quaternary vegetation history of the eastern United States, p. 381‑402. In A.R. Gillespie, S.C. Porter and B.F. Atwater, eds., 24 000, The Quaternary Period in the United States. Elsevier, Amsterdam, The Netherlands.
- Grimm, E.C. and Maher Jr., L.J., 2002. AMS Radiocarbon dating documents climate events in the upper Midwest coeval with the Bølling/Allerød and Younger Dryas episodes. Abstracts with Programs, Geological Society of America, 34 : 352.
- Hajic, E.H. and Johnson, W.H., 1989. Catastrophic flooding in the Illinois River valley; “Kankakee Torrent” revisited. Abstracts with Programs, Geological Society of America, 21 : p. 280.
- Hall, S.A., 1981. Deteriorated pollen grains and the interpretation of Quaternary pollen diagrams. Review of Paleobotany and Palynology, 2 : 193‑206.
- Hansel, A.K. and Johnson, W.H., 1992. Fluctuations of the Lake Michigan lobe during the late Wisconsin subepisode. Sveriges Geologiska Undersökning, 81 : 133‑14.
- Hansel, A.K. and Johnson, W.H., 1996. Wedron and Mason Groups: Lithostratigraphic reclassification of deposits of the Wisconsin Episode, Lake Michigan Lobe area. Illinois State Geological Survey, Bulletin 104, 116 p.
- Heusser, L., Maenza-Gmelch, T., Lowell, T. and Hinnefeld, R., 2002. Late Wisconsin periglacial environments of the southern margin of the Laurentide Ice Sheet reconstructed from pollen analyses. Journal of Quaternary Science, 17 : 773‑380.
- Iannicelli, M., 2003. Reinterpretation of the original Dekalb (sic) Mounds in Illinois. Physical Geography, 24 : 170‑182.
- Jackson, S.T., Webb, R.S., Anderson, K.H., Overpeck, J.T., Webb, T., Williams, and Hansen, B.S.C., 2000. Vegetation and environment in eastern North America during the Late Glacial Maximum. Quaternary Science Reviews, 19 : 489‑108.
- Jones, J.G., Simon, B.M. and Gardener, S., 1982. Factors affecting methanogenesis and associated anaerobic processes in the sediments of a stratified eutrophic lake. Journal of General Microbiology, 128 : 1‑11.
- Johnson, W.H. and Hansel, A.K., 1989. Age, stratigraphic position, and significance of the Lemont Drift, northeastern Illinois. Journal of Geology, 97 : 301‑318.
- Kaiser, K., 1994. Two Creeks Interstade dated through dendrochronology and AMS, Quaternary Research, 42 : 288‑298.
- Karrow, P.F., 1990. Interglacial beds at Toronto. Géographie physique et Quaternaire, 44 : 289‑297.
- Konen, M.E., Stromberg, E.T. and Curry, B.B., 2005. The De Kalb Mounds revisited. Abstracts with Programs, Geological Society of America 37 : p. 22.
- Larson, G.J., Lowell, T.V. and Ostrum, N.E., 1994. Evidence for the Two Creeks interstade in the Lake Huron Basin. Canadian Journal of Earth Sciences, 31 : 793‑797.
- Liu, C.L., Riley, K.M., and Coleman, D.D., 1986. Illinois State Geological Survey radiocarbon dates IX. Radiocarbon, 28 : 110‑122.
- Lowell, T.V., Hayward, R.K., and Denton, G.H., 1999. Role of climate oscillations in determining ice margin position: hypothesis, examples, and implications, p. 193‑203. In D.M. Mickelson and J.W. Attig, eds., Glacial Processes, Past and Present. Geological Society of America, Special Paper 337, 203 p.
- MacDonald, G.M., Beukens, R.P. and Kieser, W.E., 1991. Radiocarbon dating of limnic sediments: A comparative analysis and discussion. Ecology, 72 : 1150‑1155.
- Maher Jr., L.J., Miller, N.G., Baker, R.G., Curry, B.B. and Mickelson, D.M., 1998. Paleobiology of the sand beneath the Valders diamicton at Valders, Wisconsin. Quaternary Research, 49 : 208‑221.
- McAndrews, J.H. 1984. Pollen analysis of the 1973 ice core from Devon Island Glacier, Canada. Quaternary Research, 52 : 68‑76.
- McAndrews, J.H. and Jackson, L.J., 1988. Age and environment of the late Pleistocene mastodont and mammoth in southern Ontario. Bulletin of the Buffalo Society of Natural Sciences, 33 : 161‑172.
- Menzies, J. and Shilts, W.W., 1996. Subglacial environments, p. 15‑136. In J. Menzies, ed., Past Glacial Environments, Volume 2. Butterworth-Heinemann, Oxford, United Kindgom, 598 p.
- Mickelson, D.M., Clayton, L., Fullerton, D.S., and Borns Jr., H.W., 1983. The Late Wisconsin glacial record of the Laurentide Ice Sheet in the United States, p. 3‑37, In S. Porter and H.E. Wright Jr., eds., Late Quaternary environments of the United States, Volume 1, The Late Pleistocene. Minneapolis, University of Minnesota Press, 407 p.
- Miller, N.G. and Thompson, G.G., 1979. Boreal and western North American plants in the late Pleistocene of Vermont. Journal of the Arnold Arboretum, 60 :167‑218.
- Miller, N.G. and Spear, R.W., 1999. Late-Quaternary history of the alpine flora of the New Hampshire White Mountains. Géographie physique et Quaternaire, 53 : 137‑157.
- Morgan, A.V. and Morgan, A., 1979. Paleoentomological methods of reconstructing paleoclimate with reference to interglacial and interstadial insect faunas of southern Ontario, p. 173‑192. In W.C. Mahaney, ed., Quaternary Paleoclimate. Dowden, Hutchinson & Ross Inc., Stroudsburg, Pennsylvania, 464 p.
- Porsild, A.E., 1947. Dryas in North America. Canadian Field Naturalist, 6 : 175‑192.
- Porsild, A. E. and Cody, W. J., 1980. Vascular plants of continental Northwest Territories, Canada. National Museums of Canada, 667 p.
- Richard, P.J.H., 1979. Palaeoclimatic Significance of the late-Pleistocene and Holocene pollen record in south-central Quebec, p. 335‑360. In W.C. Mahaney, ed., Quaternary Paleoclimate. Geo Abstracts, Norwich, England, 464 p.
- Ritchie, J. C., and Harrison, S. P., 1993. Vegetation, lake levels, and climate in western Canada during the Holocene, p. 401‑414. In H.E. Wright Jr., J.E. Kutzbach, T. Webb III, W.F. Ruddiman, F.A. Street-Perrott, and P.J. Bartlein, eds., Global Climates Since the Last Glacial Maximum, University of Minnesota Press, Minneapolis, Minnesota, 569 p.
- Springer, J.W. and Flemal, R.C., 1981. Paleontological and geological results from two fossil proboscidean finds in northern Illinois. Transactions of the Illinois State Academy of Science, 74 : 87‑99.
- Staplin, F.L.,1963. Pleistocene ostracoda of Illinois. Part II. Subfamilies Cyclocyprinae, Cypridopsinae, Ilyocyprinae; Families Darwinulidae and Cytheridae. Stratigraphic ranges and assemblage patterns. Journal of Paleontology, 37 : 1164‑1203.
- Terasmae, J., Webber, P.J. and Andrews, J.T., 1966. A study of late-Quaternary plant-bearing beds in north-central Baffin Island, Canada. Arctic, 19 : 296‑318.
- Tuthill, S.L., Clayton, L., and Laird, W.M., 1964. A comparison of a fossil Pleistocene molluscan fauna from North Dakota with a recent molluscan fauna from Minnesota. American Midland Naturalist, 71 : 344‑362.
- Wang, H., Follmer, L.R. and Liu, C.L.J., 2000. Isotope evidence of paleo-ENSO cycles in the loess-paleosol record in the Central USA. Geology, 28 : 771‑774.
- Wang, H., Hughes, R.E., Steele, J.D., Lepley, S.W. and Tian, J., 2003. Correlation of climate cycles in middle Mississippi Valley loess and Greenland ice. Geology, 31 : 179‑182.
- Warner, B.G., Morgan, A.V. and Karrow, P.F., 1988. A Wisconsinan interstadial arctic flora and insect fauna from Clarksburg, southwestern Ontario, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology, 68 : 27‑47.
- Warner, B.G., Kubiw, H.J. and Karrow, P.F., 1991. Origin of a postglacial kettle-fill sequence near Georgetown, Ontario. Canadian Journal of Earth Science, 28 : 1965‑1974.
- Welsh, S.L., 1974. Anderson’s Flora of Alaska and Adjacent Parts of Canada. Bringham Young University Press, Provo, Utah, 724 p.
- West, R.G. and Petit, M.E., 2000. Plant macrofossil remains from recent sediments of Banks Island, Northwest Territories, and Bathurst Island, Nunavut, Canada, and the interpretation of Quaternary cold stage plant macrofossil assemblages. Journal of Quaternary Science, 15 : 177‑184.
- Wickham, S.S. and Johnson, W.H., 1988. Regional geology of the Tiskilwa Till Member, Wedron Formation, northeastern Illinois. Illinois State Geological Survey, Circular 543, 48 p.
- Williams, J.W., Shuman, B.N. and Webb III, T., 2001. Dissimilarity analyses of late Pleistocene and Holocene vegetation and climate in eastern North America. Ecology, 82 : 3346‑3362.
- Willman, H.B. and Payne, J.N., 1942. Geology and mineral resources of the Marseilles, Ottawa, and Streator Quadrangles. Illinois State Geological Survey, Bulletin 66, 388 p.
- Yansa, C.H., in press. The timing and nature of Late Quaternary vegetation changes in the northern Great Plains, USA and Canada: A re-assessment of the spruce phase. Quaternary Science Reviews.
- Yu, Z., 2003. Late Quaternary dynamics of tundra and forest vegetation in the southern Niagara Escarpment, Canada. New Phytologist, 157 : 365‑390.
 10.7202/032681ar
10.7202/032681arListe des figures
A
B
Study area. (A) Location of the three sublobes of the Lake Michigan lobe in northeastern Illinois, and their associated moraines (in gray). (B) Location in context of the major lobes of the Laurentide Ice Sheet in the southwestern Great Lakes region.
Région à l’étude. (A) Carte des trois sous-lobes du lobe du Lac Michigan dans le nord-est de l’Illinois et de leurs moraines respectives (en gris). (B) Carte générale des lobes majeurs de la calotte glaciaire Laurentienne dans la région du sud-ouest des Grands Lacs.
A
Marengo Phase
B
Putnam Phase
C
Livingston Phase
D
Woodstock Phase
Location of study sites and features noted in text. (A) Landforms associated with the Marengo Phase and sites with pre-glacial organics and the supraglacial ice-walled lake site (H‑3). (B) Landforms associated with the Putnam Phase (black lines) with features identified in Figure 2A (white lines). The Fox River valley is shown, although it was eroded primarily during the Woodstock Phase. (C) Landforms associated with the Livingston Phase (black lines) and older features (white lines). The sites occur on diamicton of the Yorkville Member, Lemont Formation. (D) Landforms associated with the Woodstock Phase (black lines) and older features (white lines). The sites occur on outwash associated with the Haeger Member except for the Randall Road and Sleepy Hollow sites which are associated with aggradation after the Fox River Torrent.
Localisation des sites d’étude et des structures géomorphologiques mentionnées dans le texte. (A) Morphologies associées avec la Phase de Marengo et sites de matières organiques dans les sédiments du lac de barrage glaciaire (H‑3). (B) Morphologies associées avec la phase de Putnam (trait gras) avec les structures identifiées dans la Figure 2A (lignes blanches). La vallée de la rivière Fox est indiquée ; elle a été érodée principalement pendant la Phase de Woodstock. (C) Morphologies associées avec la Phase de Livingston (lignes noires) et les morphologies plus anciennes (lignes blanches). Les sites sont associés au Membre de Yorkville, Formation de Lemont. (D) Morphologies associées avec la Phase de Woodstock et les morphologies plus anciennes (lignes blanches). Les sites sont associés avec le Membre de Haeger sauf pour les sites de Randall Road and Sleepy Hollow qui sont associés avec l’aggradation qui a eu lieu après l’événement torrentiel de la rivière Fox.
Figure 3
Stratigraphic relations between the Mason and Wedron Groups (after Hansel and Johnson, 1996; Curry et al., 1997b).
Relations stratigraphiques entre le Groupe de Mason et le Groupe de Wedron (d’après Hansel et Johnson, 1996; Curry et al., 1997b).
Figure 4
Lithologic logs of five key sites in northeastern Illinois indicating the relative abundance of ostracodes and plant macrofossils that provided the radiocarbon ages. The corresponding numbers with macrofossil symbols are the uncorrected radiocarbon ages in ka.
Relevés lithologiques de cinq sites clés dans le nord-est de l’Illinois indiquant l’abondance relative d’ostracodes et de plantes fossiles qui fournissent des âges au radiocarbone. Les nombres associés aux symboles de macrofossile sont des âges au radiocarbone non-corrigés en milliers d’années.
A
Dryas integrifolia
B
Salix herbacea
C
Vaccinium uligonosum spp. alpinium
D
Sibbaldia procumbens
E
Armeria maritima
F
Silene cf. S. acaulis
G
cf. Cardamine bulbosa
H
Ranunculus cf. R. aquatilis
I
Picea mariana
Scanning Electron Microscope (SEM) micrographs of selected plant macrofossils. Note the reference scales (white line) are of various lengths.
Sélection de photographies des plantes macrofossiles obtenues à partir d’un microscope électronique à balayage. Notez que les échelles de référence sont de longueur variable.
Figure 6
Plant macrofossil abundance diagram showing counts for both the Fox River Stone Company (FRSC) (older) and Prairie Pit #94 (PP #94) (younger) sites.
Diagramme de comptage des plantes macrofossiles pour le site Fox River Stone Company (FRSC) (plus ancien) et le site Prairie Pit #94 (PP #94) (plus jeune).
A
Limnocythere friabilis, female, left valve
B
Cytherissa lacustris, left valve
C
Limnocythere herricki, male, left valve
D
Limnocythere friabilis, female, right valve
E
Cytherissa lacustris, right valve
F
Limnocythere herricki, male, right valve
G
Heterocypris incongruens, female, left valve
H
Candona subtriangulata, female, right valve
I
Limnocythere herricki, male, right valve
Scanning Electron Microscope (SEM) micrographs of selected ostracodes.
Sélection de photographies d’ostracodes obtenues à partir d’un microscope électronique à balayage.
Figure 8
Relative abundance of ostracode valves and Pisidium clam shells from the Fox River Stone Company monolith.
Abondance relative des valves d’ostracode et de Pisidium du monolithe de Fox River Stone Company.
A
B
Time-distance diagrams for the active margin of the Harvard sublobe, northeastern Illinois. See Figure 1A for the line of section. (A) Diagram based on the radiocarbon ages in Springer and Flemal (1981), Curry et al. (1997a), Curry et al. (1999). (B) Revised time-distance diagram based on the new radiocarbon ages (Table I). We reject the ages from the Wedgewood and NIU‑123 sites (see discussion).
Diagramme de distance-temps pour la marge active du sous-lobe de Harvard, nord-est de l’Illinois. Se référer à la Figure 1A pour la localisation de la section. (A) Diagramme basé sur les âges au radiocarbone dans Springer et Flemal (1981), Curry et al. (1997a), Curry et al. (1999). (B) Diagramme de distance-temps révisé basé sur les nouveaux âges au radiocarbone (Table I). Nous excluons les âges provenant des sites de Wedgewood et de NIU‑123 (voir la discussion).
Liste des tableaux
Table I
Selected radiocarbon ages from northeastern Illinois
N.D. = not determined; N.A. = not applicable
Sites: (1) Mastodon Lake, Phillips Park, Aurora, NW NW Section 35, T. 38 N., R. 8 E.; (2) Brewster Creek, Excavation pit near boring BC‑1, Section 6, T. 40 N., R. 9 E. (Curry et al., in prep.); (3) Crystal Lake, NW Section 6, T. 43 N., R. 8 E.; (4) Sleepy Hollow meander scar, NW NW Section 28 , T. 42 N., R. 8 E.; (5) Bridge boring, Randall Road over Tyler Creek, NW Sect. 9, T. 42 N., R. 8 E.; (6) Nancy Drive, Crystal Lake, SE Section 3, T. 43 N., R. 8 E.; (7) Nelson Lake, NE Section 25, T. 39 N., R. 7 E.; (8) NIU‑123, NE SW Section 22, T. 39 N., R. 9 E. (Springer and Flemal, 1981); (9) Prairie Pit #94, NW Section 26, T. 41 N., R. 8 E.; (10) Fox River Stone Company, NW Section 4, T. 40 N., R. 8 E. (Curry et al., 1999); (11) Sleepy Hollow subdivision flood retention pond, SW Section 6, T. 42 N. R. 8 E.; (12) Ice-walled lake deposit, S 1/2 Section 34, T. 42. N., R. 6 E.; (13) Wedgewood subdivision (temporary exposure at retention pond) NE Section 12, T. 42 N., R. 7 E.; (14) Bluff City mine portal, NE SE Section 30, T. 41 N., R. 9 E.; (15) Feltes sand and gravel pit, NE Section 19, T. 39 N., R. 7 E. (Curry et al., 1999)
Radiocarbon ages <21 000 BP were calibrated using CALIB5 (http://calib.qub.ac.uk/calib/calib.html)
Radiocarbon ages >21 000 BP were calibrated using CALPAL online (http://www.calpal-online.de/); a sigma-one value is given
Table II
Difference in 14C age between ice-marginal sites high in the landscape and basal kettle dates lower in the landscape associated with the same moraine