
Résumés
Résumé
Durant ces 20 dernières années, les microbiologistes ont pris conscience que le mode de croissance bactérien utilisé en laboratoire avait ses limites. En effet, dans leur environnement naturel, les micro-organismes sont attachés à une surface, organisés en communautés structurées, et englobés dans une matrice d’exopolysaccharide. Ce mode de développement, appelé biofilm, a pris une importance toute particulière lorsqu’il a été établi qu’il était impliqué dans un grand nombre d’infections bactériennes. Pseudomonas aeruginosa est un pathogène opportuniste responsable d’infections nosocomiales et d’infections irréversibles et mortelles chez les malades souffrant de mucoviscidose. Cet organisme s’installe dans les tissus sous forme de biofilm, mais est également capable d’adhérer à des surfaces inertes. Le développement de cribles génétiques et le séquençage du génome de P. aeruginosa ont permis d’obtenir de nombreuses informations permettant de mieux comprendre ce phénomène au niveau moléculaire.
Summary
Bacterial attachment on various surfaces mostly takes place in the form of specialised bacterial communities, referred to as biofilm. The biofilm is formed through series of interactions between cells and adherence to surface, resulting in an organised structure. In this review we have been using Pseudomonas aeruginosa as a model microorganism to describe the series of events that occurred during this developmental process. P. aeruginosa is an opportunistic pathogen that has a wide variety of hosts and infectious sites. In addition to biofilm formation in certain tissues, inert surfaces, such as catheters, are also target for bacterial biofilm development. The use of convenient genetic screens has made possible the identification of numerous biofilm-defective mutants, which have been characterised further. These studies have allowed the proposal for a global model, in which key events are described for the different stages of biofilm formation. Briefly, flagellar mobility is crucial for approaching the surface, whereas type IV pili motility is preponderant for surface colonisation and microcolonies formation. These microcolonies are finally packed together and buried in an exopolysaccharide matrix to form the differentiated biofilm. It is obvious that the different stages of biofilm formation also involved perception of environmental stimuli. These stimuli, and their associated complex regulatory networks, have still to be fully characterised to understand the bacterial strategy, which initiates biofilm formation. One such regulatory system, called Quorum sensing, is one of the key player in the initial differentiation of biofilm. Finally, a better understanding, at the molecular level, of biofilm establishment and persistence should help for the design of antimicrobials that prevent bacterial infections.
Corps de l’article
La plupart des micro-organismes favorisent un mode de vie où la population bactérienne se trouve fixée sur un support (état sessile) plutôt que libre et isolée dans le milieu environnemental (état planctonique). L’attachement sur une surface est une « stratégie de survie » qui permet à la bactérie de s’installer et de coloniser un environnement. L’état planctonique pourrait se réduire au passage de la bactérie d’une surface à l’autre. Après attachement sur un support, les bactéries vont mettre en place et développer une communauté organisée à laquelle William Costerton a donné le nom de « biofilm » [1]. Jusqu’à récemment, ces biofilms, et l’activité bactérienne qui en résulte, étaient connus pour les problèmes inhérents à leur capacité de recouvrir et de corroder les canalisations ou encore les coques de bateaux. Depuis quelques années, il est apparu que leur importance dans le milieu médical est capitale, puisque 65 % des infections bactériennes chez l’homme impliquent des biofilms [2]. Les biofilms peuvent se former au niveau de cathéters ou d’implants (valves cardiaques, hanches artificielles), et attaquer des tissus corporels comme les dents, les yeux, les poumons, les oreilles ou le tractus urogénital [1]. L’étude des mécanismes moléculaires qui permettent la formation des biofilms devrait permettre de mieux maîtriser les développements pharmaceutiques et technologiques visant à prévenir ou à éradiquer leur formation.
Qu’est ce qu’un biofilm ?
Le biofilm se définit comme une population bactérienne adhérée à une surface et enrobée d’une matrice d’exopolysaccharide. L’étape initiale d’attachement fait intervenir des appendices générateurs de mouvement qui permettent d’approcher la surface à coloniser [3]. Cette approche conduit à un attachement transitoire pendant lequel la bactérie va chercher à « évaluer » la surface sur laquelle elle se trouve. Dans un deuxième temps, une association stable avec la surface ou avec d’autres micro-organismes déjà présents s’établit. Ces rassemblements de bactéries conduisent à la formation de micro-colonies dont la différenciation mène à l’élaboration du biofilm [1]. La matrice d’exopolysaccharide, essentiellement l’alginate pour Pseudomonas aeruginosa, représente quelque 85 % du volume total. Cette matrice renforce la structure du biofilm tout en lui conservant une grande plasticité. Au sein du biofilm, les micro-colonies sont séparées par des canaux aqueux qui forment un réseau de circulation permettant, d’une part, d’acheminer l’oxygène et les nutriments dans les régions enfouies du biofilm, et, d’autre part, d’évacuer les déchets. Le matériel soluble peut diffuser à travers la matrice d’exopolysaccharide et être utilisé par les bactéries. Un gradient de nutriments et d’oxygène est observable depuis le sommet du biofilm jusqu’à sa base où l’on a un micro-environnement anaérobie. Cette observation conforte l’idée selon laquelle l’état métabolique d’une bactérie à l’intérieur d’un biofilm dépend de sa localisation au sein de la structure.
Figure 1
Sélection de mutants de P. aeruginosa incapables de former un biofilm.
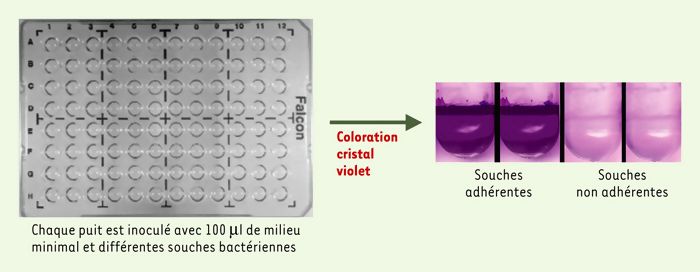
Les puits contiennent 100 μl de milieu minimal (M63, glucose 0,2 %, casamino-acides 0,5 %) et sont inoculés avec différentes souches bactériennes. Les plaques sont ensuite incubées pour des périodes de 2 à 8 heures à 30°C sans agitation. Le milieu de culture ainsi que les bactéries planctoniques sont ensuite éliminés par rinçage et la formation du biofilm bactérien est révélée par coloration au cristal violet. Pour P. aeruginosa, qui est une bactérie aérobie stricte, le biofilm se forme à l’interface air-liquide, ce qui correspond ici à la position de l’anneau violet. Dans le cas d’un mutant non adhérent, aucun anneau n’est visualisé après coloration. Ce crible, pratique et rapide, permet de tester de larges collections de mutants.
Un modèle d’étude : Pseudomonas aeruginosa
De nombreuses bactéries, importantes sur le plan médical, ont fait l’objet d’études quant à leur capacité de former des biofilms [1]. On peut citer par exemple des bactéries à Gram positif, comme Staphylococcus aureus ou Staphylococcus epidermidis, et des bactéries à Gram négatif comme Vibrio cholerae, Escherichia coli ou P. aeruginosa. Parmi celles-ci, P. aeruginosa a fait l’objet d’études assez complètes et informatives sur les mécanismes moléculaires qui interviennent lors du développement d’un biofilm.
P. aeruginosa est une bactérie à Gram négatif que l’on trouve dans de nombreux environnements comme le sol, l’eau ou la végétation. C’est également un pathogène opportuniste capable d’infecter une multitude d’hôtes comme les nématodes, les insectes, l’homme et même les plantes [4]. Chez l’homme, P. aeruginosa est la cause majeure d’infections nosocomiales chez les patients immunodéprimés. Parmi les affections les plus courantes, on peut citer les pneumonies, les infections du tractus urinaire, des brûlures, des plaies ou bien encore les septicémies. De plus, P. aeruginosa est directement responsable du taux de mortalité élevé et précoce chez les patients atteints de mucoviscidose [1]. La pathogénicité de P. aeruginosa est multifactorielle et met en jeu des adhésines permettant l’attachement, les facteurs sécrétés que sont les toxines et les enzymes [5], ou des métabolites comme le cyanide et les phénazines. L’infection chronique se caractérise par la formation du biofilm bactérien, comme c’est le cas dans le poumon des patients atteints de mucoviscidose.
L’éradication d’un biofilm bactérien pose de graves problèmes sur le plan médical. En effet, si les traitements antibiotiques classiques sont efficaces sur les bactéries planctoniques, ils s’avèrent bien moins probants sur un biofilm [6]. Lorsque l’on sait que P. aeruginosa est naturellement résistant à un certain nombre d’antibiotiques, de par la faible perméabilité de son enveloppe cellulaire mais également par la présence de nombreuses pompes d’efflux [7], qui rejettent hors de la cellule les drogues qui y pénètrent en faible quantité, on imagine la difficulté à trouver un traitement efficace pour les patients atteints de mucoviscidose. La formation du biofilm est également un facteur de résistance face à la réponse immunitaire de l’hôte [1]. La raison précise de ces phénomènes n’a pas été clairement établie et plusieurs hypothèses ont été avancées. Les bactéries au sein du biofilm sont dans un environnement pauvre en nutriments et par conséquent ont un métabolisme ralenti et une croissance lente, ce qui rend inefficace une grande majorité d’antibiotiques. Ou encore, la présence de la matrice d’exopolysaccharide ralentit la pénétration d’antibiotiques et de biocides. Ainsi, l’alginate peut capturer le peroxyde d’oxygène, les radicaux libres relargués par les macrophages ou encore certains antibiotiques.
Les facteurs de mobilité et d’adhérence
G.A. O’Toole et R. Kolter ont développé une méthode de criblage génétique qui a permis de caractériser un certain nombre de déterminants importants dans la formation de biofilm [3]. Ce criblage est fondé sur la capacité de la population bactérienne de s’attacher à la paroi des puits d’une plaque de microtitration, la présence d’un biofilm étant alors révélée par une coloration au cristal violet (Figure 1). Dans le cas de P. aeruginosa, le film est localisé à l’interface air-liquide. Ce criblage relativement simple a permis de tester de larges collections de mutants. G.A. O’Toole et R. Kolter ont ainsi mis en évidence deux classes de mutants, l’une étant défectueuse dans la mobilité flagelle, l’autre dans la biogenèse des pili de type IV [3].
Le flagelle est un appendice extracellulaire, polaire chez P. aeruginosa, constitué essentiellement par la sous-unité FliC et permettant à la bactérie de se déplacer en milieu liquide ou semi-solide. G.A. O’Toole et R. Kolter ont confirmé, en utilisant la microscopie, qu’un mutant défectueux dans la formation du flagelle n’adhère que très faiblement au support [3]. Récemment, nous avons montré que non seulement la présence du flagelle mais, également, son fonctionnement normal est important pour l’attachement au support [8]. En effet, des mutants dans les gènes motA ou motY, qui codent pour des éléments du moteur du flagelle mais dont l’inactivation n’empêche pas l’assemblage, sont incapables d’adhérer à un support. Ces observations indiquent que le flagelle est particulièrement important pour l’approche du support.
Les pili de type IV sont des structures fibrillaires présentes au pôle de certaines bactéries à Gram négatif, dont P. aeruginosa, et qui sont impliquées dans un type particulier de mobilité appelée twitching motility [9]. Ils permettent plus particulièrement des mouvements à l’interface de surfaces solides, mouvements fondés sur la capacité de rétraction de ces pili. La biosynthèse des pili de type IV fait intervenir plus d’une vingtaine de gènes, la sous-unité structurale étant codée par le gène pilA. Contrairement aux mutants dépourvus d’un flagelle fonctionnel, les mutants dont la biogenèse des pili de type IV est affectée peuvent former une monocouche cellulaire sur un support, mais sont en revanche incapables de former les micro-colonies caractéristiques [3]. Cette observation confirme que la formation des micro-colonies est bien un mécanisme d’agrégation cellulaire qui nécessite une mobilité, et non pas seulement une croissance clonale à partir d’une cellule bactérienne donnée.
Chez d’autres organismes, comme Escherichia coli, il a été montré que des appendices extracellulaires comme les fimbriae [10] ou les « curli » [11] sont également importants lors de la formation du biofilm. En utilisant le type de criblage décrit par O’Toole et Kolter, mais en modifiant légèrement les conditions de croissance, nous avons pu identifier chez P. aeruginosa de nouveaux facteurs d’attachement [8]. En effet, des mutations dans les gènes que nous avons appelés cupA (cup pour chaperone usher pathway) empêchent l’attachement au plastique. La voie chaperone usher pathway permet l’assemblage de pili de type fimbriae, qui sont essentiels à l’adhérence des E. coli uropathogènes [12]. Ce type de système n’avait jusqu’alors pas été décrit chez P. aeruginosa.
Le flagelle, les pili de type IV ou encore les Cup sont donc des appendices de surface, essentiels à la formation du biofilm, plus particulièrement dans sa phase initiale. On a vu en particulier comment le flagelle et les pili de type IV permettent l’approche et la colonisation de la surface. Dans l’étape suivante, ces structures ne sont probablement plus nécessaires et laissent la place à d’autres déterminants qui vont permettre la structuration et la différenciation en biofilm. Récemment, Peter Greenberg et ses collaborateurs [13] ont réalisé une étude visant à déterminer la variation de l’expression des gènes chez des bactéries organisées en biofilm par rapport à des bactéries planctoniques grâce à l’utilisation de puces à ADN du génome de P. aeruginosa. Cette étude montre, entre autres, que l’expression des gènes codant pour PilA, la sous-unité structurale du pilus de type IV, FliC, la sous-unité structurale du flagelle, et CupA1, une sous-unité de type fimbriae, diminue en contexte biofilm. Cela confirme bien que ces structures ne sont pas nécessaires au maintien du biofilm, une fois le développement débuté.
Les facteurs de différenciation
Le quorum sensing
Au cours de ces dernières années, un mode de régulation remarquable a été mis en évidence chez les bactéries, qui correspond à un mode de communication entre bactéries d’une même espèce. Ce système, dit quorum sensing, est considéré comme un mécanisme de régulation clé dans l’adaptation écologique et la pathogénicité de P. aeruginosa [14]. Ce système est fondé, chez les bactéries à Gram négatif, sur la production de phéromones diffusibles, des acyl-homosérine lactones (acyl-HSL), qui donnent une indication de la densité cellulaire dans un environnement donné. Les HSL produits par une bactérie donnée diffèrent par la longueur et les substitutions de leurs chaînes d’acides gras (Figure 2A). Ces HSL sont synthétisés par une HSL-synthase qui est codée par un gène de type « I ». Ces molécules diffusent à travers l’enveloppe cellulaire bactérienne et, lorsque leur concentration atteint un certain seuil, elles provoquent l’activation d’un régulateur transcriptionel de type « R » qui va alors déclencher l’expression de gènes cibles. Le gène de type « I » est un de ces gènes cibles et les molécules d’HSL sont appelées auto-inducteurs. À ce jour, trois systèmes de type quorum sensing ont été caractérisés chez P. aeruginosa, il s’agit de lasR/lasI, rhlR/rhlI et qscR [15, 16]. Ces systèmes sont connectés au sein d’une cascade et contrôlent l’expression de la quasi-totalité des gènes impliqués dans la virulence.
Figure 2
Les acyl-homosérine lactones (acyl-HSL).

A. Les deux HSL produites par P. aeruginosa diffèrent par la longueur de leur chaîne carbonée. Le 3-oxo-C12-HSL est associé au système LasR/LasI, alors que le C4-HSL fonctionne avec le système RhlR/RhlI. B. Inhibition du quorum sensing par l’algue marine Delisea pulchra. Chromobacterium violaceum produit, entre autres, en réponse au quorum sensing (C6-HSL), un pigment violet et peut donc être utilisé comme senseur de la production d’HSL. La présence de D. pulchra, qui produit les furanones, inhibe la production de ce pigment. Les furanones interfèrent avec le fonctionnement du quorum sensing chez C. violaceum, et en particulier avec le fonctionnement du C6-HSL.
Une observation cruciale et totalement originale a été réalisée par Davies et al. [17]. Ces auteurs rapportent que la structure d’un biofilm, formé par une souche défectueuse dans la production d’HSL (lasI), est plate et non différenciée en raison de l’absence de formation de micro-colonies. En revanche, les bactéries sont plus nombreuses, plus compactes et distribuées de façon uniforme. Ce type de structure est sensible à un biocide comme le SDS. La variation d’expression des gènes liés au quorum sensing semble donc être cruciale pour la différenciation en une structure de type biofilm.
Au début des années 1990, Peter Steinberg s’est étonné de voir la capacité de l’algue marine Delisea pulchra à rester libre de toute colonisation dans des eaux pourtant infestées de bactéries. Stefan Kjelleberg et al. ont pu démontrer que la molécule, appelée furanone, qui prévient la contamination de l’algue a une structure très proche des HSL [18] (Figure 2), et se comporte comme un antagoniste des HSL. La conception de divers inhibiteurs de ce type pourrait faire l’objet de la synthèse de drogues anti-biofilm. Le fait de bloquer la formation du biofilm, ou bien de le démanteler, peut certainement aider à traiter efficacement les infections bactériennes.
L’alginate
Il est connu depuis de nombreuses années que les isolats pulmonaires de P. aeruginosa sont fortement muqueux. La mucosité est due à la production d’alginate, un exopolysaccharide produit par P. aeruginosa. Contrairement aux flagelles et aux pili, l’expression de algC est augmentée d’un facteur quatre en biofilm [19]. Il est également intéressant de constater que le facteur sigma alternatif σ22, ou AlgT, contrôle positivement la production d’alginate alors qu’il contrôle négativement la synthèse du flagelle [19]. Dans le cas d’E. coli, c’est l’acide colanique qui constitue l’enveloppe muqueuse du biofilm et dont la production est augmentée lors de l’attachement. Alors qu’une souche non muqueuse peut former un biofilm d’une épaisseur de 6 μm, une souche muqueuse peut donner un biofilm ayant une épaisseur moyenne de 40 μm. L’enrobement des bactéries par une couche d’alginate a un rôle non seulement structural, mais également protecteur. Il a été montré par exemple que des souches non muqueuses forment des biofilms qui ont une résistance beaucoup plus faible envers la tobramycine, l’un des antibiotiques les plus utilisés pour lutter contre les infections par P. aeruginosa.
Les lipopolysaccharides (LPS)
Ils forment le feuillet externe de l’enveloppe bactérienne des bactéries à Gram négatif, et semblent également jouer un rôle particulier dans la formation du biofilm. P. aeruginosa peut posséder deux types de LPS. Le LPS de type B, de haut poids moléculaire, et le LPS de type A, qui ne possède que le core-lipid A contenant les chaînes d’acides gras, et est donc un LPS de petit poids moléculaire présentant une surface plus hydrophobe. La composition en LPS, et la variation du niveau des bandes A ou B, peuvent être importantes pour l’attachement sur différents types de surface selon qu’elle est hydrophile ou hydrophobe [20].
Figure 3
Biofilm bactérien mixte.

Les bactéries fluorescentes vertes ou rouges sont visualisées par l’utilisation d’un microscope confocal (confocal scanning laser microscopy, CSLM). Les bactéries présentes dans le biofilm sont P. putida OUS82 (marquage Gfp) et Pseudomonas sp. B13 (marquage dsRed). On distingue clairement les micro-colonies de chacune des espèces (rouge ou verte). Cependant, dans chaque micro-colonie, on voit également quelques bactéries de l’autre espèce.
Autres facteurs
La liste des déterminants impliqués dans la formation du biofilm est certainement loin d’être exhaustive, notamment parce que la formation de ces structures peut répondre à de nombreux facteurs environnementaux et donc être spécifique dans des conditions de croissance déterminées. On sait maintenant que Crc (catabolite repression protein) de P. aeruginosa, qui réprime le métabolisme des hydrates de carbone en présence d’intermédiaires du cycle de Krebs (pyruvate, succinate), semble appartenir à une voie de signalisation qui contrôle la formation du biofilm. Celle-ci est en effet déficiente chez les mutants crc [21]. De façon assez spectaculaire, la mutation crc aboutit, entre autres, à un défaut de formation des pili de type IV qui peut expliquer l’incapacité à développer un biofilm.
La mise en évidence de réseaux de régulation complexes impliqués dans l’attachement des bactéries à une surface, et dans la différenciation en biofilm, pourrait contribuer à l’identification de nouveaux effecteurs essentiels à ces processus. On a évoqué le cas du quorum sensing, de Crc, mais d’autres études ont également montré que GacA, le régulateur de réponse du couple GacA/GacS, un système à deux composants, influe également sur la formation du biofilm [22]. Chez E. coli, les systèmes à deux composants EnvZ/OmpR et CpxA/CpxR, permettent de percevoir des stress au niveau de l’enveloppe bactérienne, et seraient impliqués conjointement dans la formation du biofilm [11, 23].
La variation de phase est une stratégie qui permet à la bactérie de s’adapter rapidement à des variations de conditions environnementales. Cette variation est liée à une réorganisation génétique, qui va allumer ou éteindre l’expression d’une série de gènes auxquels sont associés divers traits phénotypiques. Chez les bactéries à Gram négatif, l’expression de nombreuses structures de surface comme les fimbriae sont soumises à cette variation [12]. Récemment, Déziel et al. [24] ont mis en évidence l’émergence spontanée de variants de P. aeruginosa formant des colonies rugueuses et de petite taille lorsque la bactérie est cultivée dans un milieu statique favorable à la formation d’un biofilm. Les bactéries formant ces colonies ont, à leur surface, d’abondants pili de type IV mais apparaissent dépourvues de twitching motility aussi bien que de mobilité flagelle. Cependant, contrairement à la forme parentale, ces variants ont une forte capacité d’adhérence. Selon les conditions de culture, les bactéries de chaque type (parentale ou variant) passent d’un phénotype à l’autre avec une fréquence assez élevée. Il semble que la diversité phénotypique déterminée par la variation de phase prenne soin de maintenir, dans la population, des bactéries qui soient parfaitement adaptées à la formation de biofilm, formation qui est alors déclenchée dès que les conditions environnementales s’y prêtent.
Le biofilm, une communauté bactérienne multi-espèces
Si nous avons évoqué, jusqu’à présent, la formation du biofilm en utilisant P. aeruginosa comme modèle, il faut souligner qu’à l’état naturel les biofilms sont des communautés bactériennes au sein desquelles on peut trouver de nombreuses espèces différentes. Dans ces communautés, le transfert horizontal de gènes est favorisé et les bactéries échangent leur matériel génétique à des fréquences très élevées. Récemment, il a été proposé que la formation des pili conjugatifs était également un facteur déterminant pour la formation du biofilm proprement dit [25].
Figure 4
Modèle de formation d’un biofilm par Pseudomonas aeruginosa.
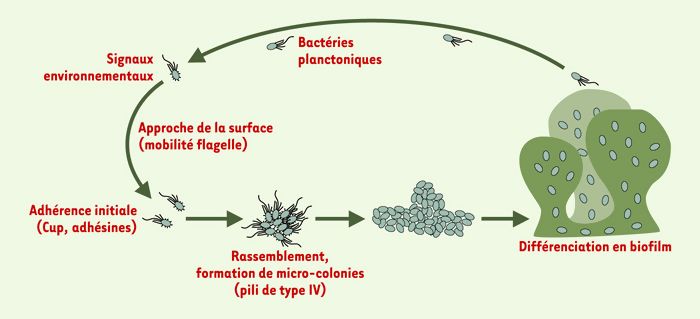
Les bactéries planctoniques vont s’attacher sur une surface en réponse à un certain nombre de stimulus environnementaux et nutritionnels. L’approche de cette surface nécessite la mobilité flagelle. L’attachement initial sur la surface fait intervenir un certain nombre d’adhésines dont les structures de type Cup nouvellement identifiées chez P. aeruginosa. La colonisation de la surface et le rassemblement en micro-colonies font intervenir un autre appendice de mobilité, les pili de type IV. La structuration des micro-colonies en biofilm mature implique l’expression de certains gènes liés au quorum sensing, la répression de l’expression de fliC, pilA et CupA1 et la production d’exopolysaccharide qui englobe les bactéries, l’alginate chez P. aeruginosa.
Les conditions qui permettent la formation d’un biofilm mixte ne sont pas clairement établies. S. Molin et al. [26] ont suggéré que la mixité des populations est en partie dépendante de la nourriture. Ainsi, en présence de chlorobiphényl comme source de carbone, Pseudomonas putida peut se nourrir des métabolites produits par Burkholderia, et les deux espèces forment un biofilm mixte. En revanche, en présence de citrate, utilisable par les deux espèces, chacune formera son propre biofilm avec une structure radicalement différente. La localisation des différentes espèces et la mixité des biofilms peuvent être observées grâce à l’utilisation de techniques de fluorescence. Les sondes phylogénétiques sont des oligonucléotides dérivés des ARN ribosomiques 16S et 23S couplés à des molécules fluorescentes [27]. Les bactéries peuvent également être étiquetées par l’introduction des gènes codant pour la GFP (green fluorescent protein) ou la DsRed (Discosomasp. red fluorescent protein), sous le contrôle d’un promoteur constitutif (Figure 3). L’expression de ces protéines fluorescentes va permettre de pister une espèce donnée dans un environnement complexe. Ainsi, il n’est pas rare d’observer à l’intérieur du biofilm des mouvements de bactéries passant d’une micro-colonie à l’autre [28]. Ces observations révèlent, qu’à l’intérieur même du biofilm, chaque micro-colonie est une petite communauté en soi.
Conclusions
Cet article n’est certes pas exhaustif et ne tend pas à énumérer tous les gènes ayant une influence sur la formation d’un biofilm. Nous avons souhaité exposer certains principes généraux qui permettent d’appréhender comment des bactéries isolées peuvent se regrouper et s’organiser en structures complexes et différenciées. À partir de là se dégage un modèle généralement admis mais, qui, il faut bien le reconnaître, comporte de nombreuses faiblesses (Figure 4). Les différents éléments qui sont décrits dans cet article ne sont pas nécessairement tous intégrés dans la succession d’événements qui conduit à l’élaboration du biofilm. Il est aussi probable qu’il n’y ait pas de système universel de formation d’un biofilm, et ce qui est vrai pour P. aeruginosa ne l’est pas nécessairement pour d’autres organismes. En effet, les voies métaboliques qui déclenchent la formation du biofilm, et les facteurs d’adhérence impliqués, dépendent de l’organisme concerné et de l’environnement dans lequel il forme le biofilm. Néanmoins, tous ces déterminants sont autant de cibles pour développer des traitements préventifs de la formation de biofilm. L’exemple des furanones démontre que le principe est valable même si ces molécules, très mutagènes et toxiques, ne sauraient être utilisées chez l’homme.
Parties annexes
Remerciements
Nous remercions Michael Givskov pour nous avoir procuré les photographies de biofilm en microscopie confocale, ainsi que la photo de l’algue marine Delisea pulchra. Nos travaux sont soutenus par Vaincre La Mucoviscidose (VLM), le Programme Recherche Fondamentale Microbiologie Maladies Infectieuses et Parasitaires (PRFMMIP), la Région Provence-Alpes-Côte d’Azur, ainsi que par la Communauté européenne (Contrat QLK2-CT-2001-01339).
Références
- 1. Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms : a common cause of persistent infections. Science 1999 ; 284 : 1318-22.
- 2. Chicurel M. Bacterial biofilms and infections. Slimebusters. Nature 2000 ; 408 : 284-6.
- 3. O’Toole GA, Kolter R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol Microbiol 1998 ; 30 : 295-304.
- 4. Rahme LG, Ausubel FM, Cao H, et al. Plants and animals share functionally common bacterial virulence factors. Proc Natl Acad Sci USA 2000 ; 97 : 8815-21.
- 5. Filloux A, Michel G, Bally M. GSP-dependent protein secretion in Gram-negative bacteria : the Xcp system of Pseudomonas aeruginosa. Microbiol Rev 1998 ; 22 : 177-98.
- 6. Brooun A, Liu S, Lewis K. A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob Agents Chemother 2000 ; 44 : 640-6.
- 7. Poole K. Multidrug efflux pumps and antimicrobial resistance in Pseudomonas aeruginosa and related organisms. J Mol Microbiol Biotechnol 2001 ; 3 : 255-64.
- 8. Vallet I, Olson JW, Lory S, Lazdunski A, Filloux A. The chaperone/usher pathways of Pseudomonas aeruginosa : identification of fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc Natl Acad Sci USA 2001 ; 98 : 6911-6.
- 9. Wall D, Kaiser D. Type IV pili and cell motility. Mol Microbiol 1999 ; 32 : 1-10.
- 10. Pratt LA, Kolter R. Genetic analysis of Escherichia coli biofilm formation : roles of flagella, motility, chemotaxis and type I pili. Mol Microbiol 1998 ; 30 : 285-93.
- 11. Prigent-Combaret C, Brombacher E, Vidal O, et al. Complex regulatory network controls initial adhesion and biofilm formation in Escherichia colivia regulation of the csgD gene. J Bacteriol 2001 ; 183 : 7213-23.
- 12. Sauer FG, Barnhart M, Choudhury D, Knight SD, Waksman G, Hultgren SJ. Chaperone-assisted pilus assembly and bacterial attachment. Curr Opin Struct Biol 2000 ; 10 : 548-56.
- 13. Whiteley M, Bangera MG, Bumgarner RE, et al. Gene expression in Pseudomonas aeruginosa biofilms. Nature 2001 ; 413 : 860-4.
- 14. Parsek MR, Greenberg EP. Acyl-homoserine lactone quorum sensing in Gram-negative bacteria : a signaling mechanism involved in associations with higher organisms. Proc Natl Acad Sci USA 2000 ; 97 : 8789-93.
- 15. Latifi A, Foglino M, Tanaka K, Williams P, Lazdunski A. A hierarchical quorum-sensing cascade in Pseudomonas aeruginosa links the transcriptional activators LasR and RhlR (VsmR) to expression of the stationary-phase sigma factor RpoS. Mol Microbiol 1996 ; 21 : 1137-46.
- 16. Chugani SA, Whiteley M, Lee KM, D’Argenio D, Manoil C, Greenberg EP. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc Natl Acad Sci USA 2001 ; 98 : 2752-7.
- 17. Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 1998 ; 280 : 295-8.
- 18. Manefield M, de Nys R, Kumar N, et al. Evidence that halogenated furanones from Delisea pulchra inhibit acylated homoserine lactone (AHL)-mediated gene expression by displacing the AHL signal from its receptor protein. Microbiology 1999 ; 145 : 283-91.
- 19. Garett ES, Perlegas D, Wozniak, DJ. Negative control of flagellum synthesis in Pseudomonas aeruginosa is modulated by the alternative sigma factor AlgT (AlgU). J Bacteriol 1999 ; 181 : 7401-4
- 20. Makin SA, Beveridge TJ. The influence of A-band and B-band lipopolysaccharide on the surface characteristics and adhesion of Pseudomonas aeruginosa to surfaces. Microbiology 1996 ; 142 : 299-307.
- 21. O’Toole GA, Gibbs KA, Hager PW, Phibbs PV, Kolter R. The global carbon metabolism regulator Crc is a component of a signal transducion pathway required for biofilm development by Pseudomonas aeruginosa. J Bacteriol 2000 ; 182 : 425-31
- 22. Parkins MD, Ceri H, Storey DG. Pseudomonas aeruginosa GacA, a factor in multihost virulence, is also essential for biofilm formation. Mol Microbiol 2001 ; 40 : 1215-26.
- 23. Dorel C, Vidal O, Prigent-Combaret C, Vallet I, Lejeune P. Involvement of the Cpx signal transduction pathway of E. coli in biofilm formation. FEMS Microbiol Lett 1999 ; 178 : 169-75.
- 24. Déziel E, Comeau Y, Villemur R. Initiation of biofilm formation by Pseudomonas aeruginosa 57RP correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming, swarming, and twitching motilitie. J Bacteriol 2001 ; 183 : 1195-204.
- 25. Ghigo JM. Natural conjugative plasmids induce bacterial biofilm development. Nature 2001 ; 412 : 442-5.
- 26. Nielsen AT, Tolker-Nielsen T, Barken KB, Molin S. Role of commensal relationships on the spatial structure of a surface-attached microbial consortium. Environ Microbiol 2000 ; 2 : 59-68.
- 27. Fuchs BM, Wallner G, Beisker W, Schwippl I, Ludwig W, Amann R. Flow cytometric analysis of the in situ accessibility of Escherichia coli 16S rRNA for fluorescently labeled oligonucleotide probes. Appl Environ Microbiol 1998 ; 64 : 4973-82.
- 28. Tolker-Nielsen T, Brinch UC, Ragas PC, Andersen JB, Jacobsen CS, Molin S. Development and dynamics of Pseudomonas sp. biofilms. J Bacteriol 2000 ; 182 : 6482-9.
Liste des figures
Figure 1
Sélection de mutants de P. aeruginosa incapables de former un biofilm.
Les puits contiennent 100 μl de milieu minimal (M63, glucose 0,2 %, casamino-acides 0,5 %) et sont inoculés avec différentes souches bactériennes. Les plaques sont ensuite incubées pour des périodes de 2 à 8 heures à 30°C sans agitation. Le milieu de culture ainsi que les bactéries planctoniques sont ensuite éliminés par rinçage et la formation du biofilm bactérien est révélée par coloration au cristal violet. Pour P. aeruginosa, qui est une bactérie aérobie stricte, le biofilm se forme à l’interface air-liquide, ce qui correspond ici à la position de l’anneau violet. Dans le cas d’un mutant non adhérent, aucun anneau n’est visualisé après coloration. Ce crible, pratique et rapide, permet de tester de larges collections de mutants.
Figure 2
Les acyl-homosérine lactones (acyl-HSL).
A. Les deux HSL produites par P. aeruginosa diffèrent par la longueur de leur chaîne carbonée. Le 3-oxo-C12-HSL est associé au système LasR/LasI, alors que le C4-HSL fonctionne avec le système RhlR/RhlI. B. Inhibition du quorum sensing par l’algue marine Delisea pulchra. Chromobacterium violaceum produit, entre autres, en réponse au quorum sensing (C6-HSL), un pigment violet et peut donc être utilisé comme senseur de la production d’HSL. La présence de D. pulchra, qui produit les furanones, inhibe la production de ce pigment. Les furanones interfèrent avec le fonctionnement du quorum sensing chez C. violaceum, et en particulier avec le fonctionnement du C6-HSL.
Figure 3
Biofilm bactérien mixte.
Les bactéries fluorescentes vertes ou rouges sont visualisées par l’utilisation d’un microscope confocal (confocal scanning laser microscopy, CSLM). Les bactéries présentes dans le biofilm sont P. putida OUS82 (marquage Gfp) et Pseudomonas sp. B13 (marquage dsRed). On distingue clairement les micro-colonies de chacune des espèces (rouge ou verte). Cependant, dans chaque micro-colonie, on voit également quelques bactéries de l’autre espèce.
Figure 4
Modèle de formation d’un biofilm par Pseudomonas aeruginosa.
Les bactéries planctoniques vont s’attacher sur une surface en réponse à un certain nombre de stimulus environnementaux et nutritionnels. L’approche de cette surface nécessite la mobilité flagelle. L’attachement initial sur la surface fait intervenir un certain nombre d’adhésines dont les structures de type Cup nouvellement identifiées chez P. aeruginosa. La colonisation de la surface et le rassemblement en micro-colonies font intervenir un autre appendice de mobilité, les pili de type IV. La structuration des micro-colonies en biofilm mature implique l’expression de certains gènes liés au quorum sensing, la répression de l’expression de fliC, pilA et CupA1 et la production d’exopolysaccharide qui englobe les bactéries, l’alginate chez P. aeruginosa.