Article body
La séquence complète d’un génome eucaryote fut obtenue pour la première fois en 1996 chez la levure de boulanger Saccharomyces cerevisiae [1]. Il apparut alors que la moitié des gènes avaient échappé aux cribles de la mutagenèse classique et que, pour bon nombre d’entre eux, leur fonction ne pouvait être prédite par l’analyse de leur séquence. Des études d’analyse fonctionnelle furent alors développées chez S. cerevisiae, comme le projet EUROFAN, financé par la Commission européenne, qui conduisit au clonage et à la délétion individuelle de 700 gènes de S.cerevisiae de fonction totalement inconnue (http://mips.gsf.de/proj/eurofan/index.html). Récemment, un consortium américano-européen a construit 5916 mutants délétés, soit une collection presque complète du génome (96,5 % des phases ouvertes de lecture annotées [ORF, open reading frame] de S. cerevisiae) [2-3]. Chaque gène a été remplacé par une cassette de délétion contenant le gène bactérien KanR responsable chez la levure de la résistance à la généticine [4], et de part et d’autre de celui-ci, deux séquences de 20 nucléotides distinctes et différentes pour chaque délétion (Figure 1). Ces séquences, qui constituent des code-barres caractérisant spécifiquement chaque gène délété, permettent d’analyser en parallèle un mélange de souches délétées. La collection de souches délétées est cultivée dans un milieu donné et des échantillons cellulaires sont prélevés au cours de la croissance. L’ADN des cassettes de délétion est extrait et amplifié par PCR grâce à deux oligonucléotides communs à toutes les cassettes de délétion, puis il est hybridé à des puces à ADN Affymétrix® correspondant aux 11832 code-barres des cassettes de délétion. Dans un mélange de souches, l’abondance d’une souche particulière, quantifiée par l’intensité du signal obtenu sur les puces à ADN, permet d’estimer son aptitude à la croissance et d’en déduire l’avantage ou le désavantage sélectif conféré par une délétion donnée dans des conditions de culture définies (Figure 2). Ainsi, si un gène est nécessaire à la croissance dans des conditions données, l’intensité du signal correspondant au code-barre de sa délétion diminuera rapidement au cours de la croissance dans ces conditions.
Figure 1
Cassette utilisée pour la délétion systématique des gènes de Saccharomyces cerevisiae.

La cassette contient le gène KanRsous le contrôle d’un promoteur reconnu par S. cerevisiae, deux séquences « code-barres » Tag1 et Tag2 spécifiques de chaque délétion, les séquences U1 et U2 communes à toutes les cassettes qui sont utilisées pour l’amplification simultanée de ces dernières, et 40 bases homologues au génome de levure, de part et d’autre du gène à déléter. Après transformation de la levure, la cassette se substitue au gène par recombinaison homologue.
Figure 2
Profil de croissance d’une souche délétée de la classe III.
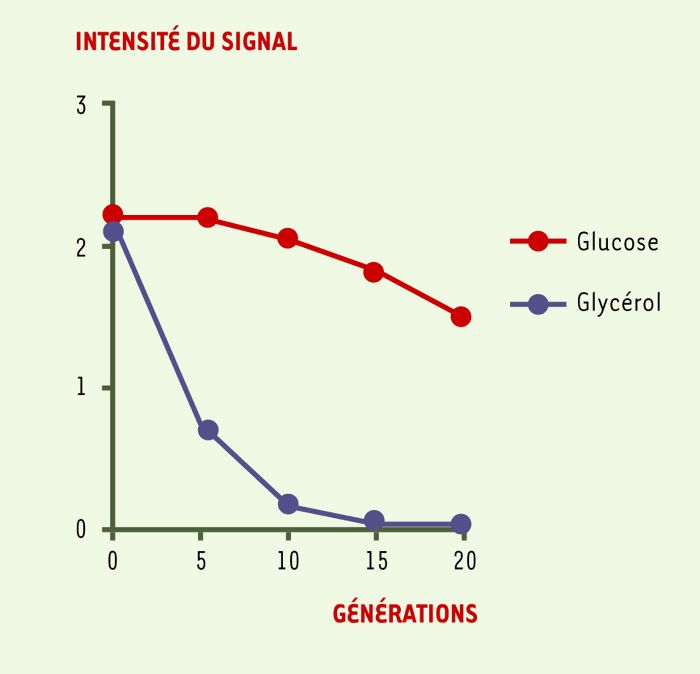
L’intensité du signal détecté sur les puces à ADN diminue plus lentement en présence de glucose (conditions non respiratoires ou fermentescibles) qu’en présence de glycérol (conditions respiratoires obligatoires ou non fermentescibles), situation au cours de laquelle la croissance de la souche s’arrête très rapidement.
Cette approche a été utilisée pour identifier les gènes de levure impliqués dans la respiration mitochondriale. L’ADN mitochondrial ne code que pour un petit nombre des polypeptides des complexes de la chaîne respiratoire et de l’ATP-synthase, la grande majorité des protéines mitochondriales étant codées par l’ADN nucléaire et importées dans la mitochondrie. On estime qu’à peine 60 % des protéines mitochondriales ont été identifiées. Quatre mille sept cent-six souches homozygotes délétées ont été cultivées sur 9 milieux discriminants pour la fonction respiratoire, contenant, par exemple, une source carbonée fermentescible comme le glucose, ou non fermentescible comme le glycérol, ou bien encore un agent découplant tel que le dinitrophénol qui perméabilise la membrane mitochondriale aux protons (Figure 2). La validité de la méthode a été vérifiée sur les 425 ORF mitochondriales de levure déjà connues, dont il a fallu soustraire 72 ORF non analysables pour diverses raisons (délétion létale ou impossible à réaliser, signal trop faible pour l’analyse sur puce à ADN). Sur les 353 ORF restantes qui produisent un signal quantifiable, 201 mutants produisent un signal plus faible en milieu non fermentescible qu’en milieu fermentescible et sont donc spécifiquement affectés dans la respiration. Dans un second temps, les 4706 souches délétées ont été classées en quatre catégories d’après leur profil en milieu respiratoire, la classe III (466 gènes) correspondant aux souches spécifiquement affectées en présence de glycérol (conditions respiratoires). Cette classification a permis de découvrir 161 nouvelles protéines, de localisation subcellulaire et de fonction inconnues, impliquées dans la respiration mitochondriale. La moitié d’entre elles contiennent un signal d’adressage à la mitochondrie, ou sont spécifiquement homologues à des protéines de Rickettsia prowazekii, considérée comme la bactérie la plus proche de la mitochondrie. Par ailleurs, 104 protéines non mitochondriales, ayant des fonctions variées au sein de la cellule, affectent également la respiration. L’identification de telles protéines est particulièrement intéressante pour comprendre comment les mitochondries sont intégrées dans les réseaux métaboliques intracellulaires.
Les gènes assurant les fonctions de base de la cellule ont été conservés au cours de l’évolution chez l’ensemble des eucaryotes, et la levure s’est révélée un outil particulièrement utile dans l’identification des gènes nucléaires impliqués dans des maladies mitochondriales chez l’homme [5]. En effet, lorsqu’un gène impliqué dans une maladie génétique est cartographié sur un chromosome, il est souvent utile de rechercher son homologue chez la levure. Dans le cas des gènes impliqués dans la respiration, 255 orthologues humains des gènes de levure de la classe III ont été identifiés, dont 21 sont connus pour leur implication dans des maladies mitochondriales. Pour sept maladies mitochondriales dont les gènes ont été cartographiés approximativement dans une région déterminée d’un chromosome, onze nouveaux gènes candidats ont été proposés, du fait de leur présence dans la région génomique cartographiée et de leur homologie avec les gènes de classe III nouvellement identifiés.
Les banques de génomique fonctionnelle mitochondriale dont nous déplorions l’absence récemment [5] ont maintenant été construites. Elles présentent cependant quelques imperfections. Les mutants de délétion ont logiquement été réalisés dans la souche de S. cerevisiae utilisée pour le séquençage du génome: malheureusement, cette souche de laboratoire contient déjà des mutations qui affectent la respiration et ne représente donc pas le modèle idéal pour l’étude de la respiration. De plus, un certain nombre de gènes ont échappé à cette analyse. Ce sont, d’une part, les gènes essentiels et, d’autre part, les gènes redondants dont le phénotype de déficience ne pourrait être décelé que par des délétions simultanées. Enfin, certaines délétions sont associées à de l’aneuploïdie qui pourrait masquer certains phénotypes [6]. De façon inattendue, certaines délétions de gènes ayant une fonction connue et apparemment non liée aux fonctions mitochondriales sont défavorables à la croissance cellulaire en milieu respiratoire et sont ainsi susceptibles d’éclairer sous un jour nouveau les relations de la mitochondrie avec les autres compartiments de la cellule. Cependant, il convient de vérifier que le phénotype mitochondrial observé ne résulte pas de mutations secondaires produites dans le génome lors de la transformation de la levure avec la cassette de délétion [6]. Enfin, le nombre de gènes considérés comme nouveaux nous semble surestimé, certains d’entre eux ayant déjà fait l’objet de publications.
L’identification de nouvelles protéines mitochondriales a également progressé par d’autres approches, comme l’analyse par spectrométrie de masse de fragments peptidiques provenant de mitochondries de levure purifiées [7]. Par ailleurs, l’analyse des profils d’expression du génome de S. cerevisiae dans 300 conditions différentes a permis de classer les gènes de levure en classes fonctionnelles [8], et en particulier d’identifier de nouvelles protéines jouant un rôle dans la mitochondrie. Cette approche expérimentale est cependant beaucoup plus lourde, et l’accès public aux résultats malaisé. Au contraire, l’étude fonctionnelle des souches délétées de S. cerevisiae, favorisée par le libre accès aux résultats obtenus pour chaque souche et aux listes des gènes de la classe III et de ses homologues humains (http://www-deletion.stanford.edu/YDPM/YDPM_index.html), constitue une source d’une richesse inestimable pour les recherches futures des fonctions de la mitochondrie chez la levure et chez l’homme, et en particulier pour la recherche des gènes responsables des maladies mitochondriales [9].
Appendices
Références
- 1. Goffeau A, Barrell BG, Bussey H, et al. Life with 6000 genes. Science 1996; 274: 563-7.
- 2. Winzeler EA, Shoemaker DD, Astromoff A, et al. Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 1999; 285: 901-6
- 3. Giaver G. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002; 418: 387-91.
- 4. Wach A, Brachat A, Pohlmann R, Philippsen P. New heterologous modules for classical or PCR-based gene disruptions in Saccharomyces cerevisiae. Yeast 1994; 10: 1793-808
- 5. Foury F, Kucej M. Yeast mitochondrial biogenesis: a model system for humans? Curr Opin Chem Biol 2002; 6: 106-11.
- 6. Grunenfelder B, Winzeler EA. Treasures and traps in genome-wide data sets: case examples from yeast. Nat Rev Genet 2002; 3: 653-61.
- 7. Pflieger D, Le Caer JP, Lemaire C, Bernard BA, Dujardin G, Rossier J. Systematic identification of mitochondrial proteins by LC-MS/MS. Anal Chem 2002; 74: 2400-6
- 8. Hughes TR, Marton MJ, Jones AR, et al. Functional discovery via a compendium of expression profiles. Cell 2000; 102: 109-26.
- 9. Steinmetz LM, Scharfe C, Deutschbauer AM, et al. Systematic screen for human disease genes in yeast. Nat Genet 2002; 31: 400-4.
List of figures
Figure 1
Cassette utilisée pour la délétion systématique des gènes de Saccharomyces cerevisiae.
La cassette contient le gène KanRsous le contrôle d’un promoteur reconnu par S. cerevisiae, deux séquences « code-barres » Tag1 et Tag2 spécifiques de chaque délétion, les séquences U1 et U2 communes à toutes les cassettes qui sont utilisées pour l’amplification simultanée de ces dernières, et 40 bases homologues au génome de levure, de part et d’autre du gène à déléter. Après transformation de la levure, la cassette se substitue au gène par recombinaison homologue.
Figure 2
Profil de croissance d’une souche délétée de la classe III.
L’intensité du signal détecté sur les puces à ADN diminue plus lentement en présence de glucose (conditions non respiratoires ou fermentescibles) qu’en présence de glycérol (conditions respiratoires obligatoires ou non fermentescibles), situation au cours de laquelle la croissance de la souche s’arrête très rapidement.