Abstracts
Résumé
Au Québec, comme dans la plus grande partie des populations mondiales, la mobilité interrégionale vers les zones urbaines et leur périphérie a gagné en importance au cours du 20e siècle. Ce travail vise à estimer, au moyen d’une approche généalogique, les effets de ces mouvements migratoires sur la variabilité du pool génique de la région de Lanaudière, située au nord-est de Montréal. Pour ce faire, la région de Lanaudière a été divisée en deux sous-régions, soit le nord et le sud. Quatre cents ascendances généalogiques d’individus mariés dans chacune des sous-régions durant deux périodes (1945-55 et 1985-95) ont été reconstruites à l’aide du fichier de population BALSAC. Pour la période 1945-55, les indices généalogiques des deux sous-régions sont très similaires, alors que pour 1985-95, ceux-ci indiquent une plus grande hétérogénéité au sud de la région, ce qui est cohérent avec le récent boom démographique enregistré dans cette partie du territoire. En outre, on a observé une réduction marquée des coefficients d’apparentement et de consanguinité et un changement des trajectoires migratoires interrégionales au cours de cette période de cinquante ans. Ces événements peuvent être clairement associés aux migrations récentes vers les zones urbaines.
Abstract
Interregional mobility towards urban areas and their periphery has become increasingly important throughout the 20th century. The effects of these migratory movements on the variability of the Lanaudiere gene pool were estimated through the use of a genealogical approach. The region of Lanaudiere was divided into two subregions, namely north and south. Four hundred ascending genealogies of individuals married within each subregion during two periods (1945-55 and 1985-95) were reconstructed using the BALSAC database. For the 1945-55 period, genealogical indices of the two subregions are quite similar, while they display much more heterogeneity in the southern part of the region in 1985-95, which is consistent with the recent demographic boom in this area. Overall, a marked reduction of kinship and consanguinity levels in Lanaudiere is observed in this fifty-year period, which can be clearly associated with changing interregional migratory trajectories toward urban areas.
Article body
Le 20e siècle a été témoin de changements démographiques majeurs. À l’échelle mondiale, la proportion de la population urbaine est passée d’environ 15 % à 50 %. La progression de l’urbanisation a été soutenue par une croissance démographique importante et une augmentation de la mobilité vers les zones urbaines et leur périphérie (Clark, 2003). Il y a plus de vingt ans, certains auteurs ont suggéré que de tels bouleversements démographiques allaient modifier la structure génétique des populations humaines, et qu’il serait urgent de documenter la diversité génétique présente au sein des populations mondiales (Cavalli-Sforza etal., 1991). Les conséquences génétiques des mouvements migratoires vers les zones urbaines ont des retombées potentielles pour la santé des individus en raison de l’augmentation de l’hétérogénéité des populations urbaines (Campbell et al., 2007; Rudan et al., 2008) et pour les études de cartographie génétique, du fait, notamment, de la réduction de la stratification des sous-populations (Helgason et al., 2005; Vitart et al., 2005). Pourtant, jusqu’à aujourd’hui, les modifications de la structure génétique des populations causées par ces mouvements migratoires n’ont été évaluées empiriquement qu’en de rares occasions (Helgason et al., 2005; Ashrafian-Bonab et al., 2007).
L’impact des migrations sur la structure génétique des populations d’accueil dépend du nombre relatif de migrants et de leur diversité, mais aussi des modalités de leur éventuelle intégration. En effet, l’apport génétique des migrations à la population d’accueil se vérifie seulement si les migrants contribuent à la génération suivante. Ainsi, pour observer directement le flux génétique, il ne faut pas seulement suivre les trajectoires des migrants, mais aussi leur succès reproducteur (Heyer et Cazes, 1999). Il est possible d’évaluer l’importance de ces variables en étudiant les liens ancestraux unissant les individus contemporains dans les populations où les registres civils permettent la reconstruction de généalogies extensives et profondes. Par conséquent, en plus de permettre d’estimer la diversité génétique d’une population, les généalogies procurent des données empiriques sur l’évolution des phénomènes démographiques qui façonnent cette diversité. À titre d’exemple, des travaux sur l’apport des immigrants fondateurs ont été réalisés dans les populations de Tristan da Cunha (Roberts, 1968), de l’Islande (Helgason, 2003), et au Québec, dans les populations du Québec ancien (Charbonneau et al., 1987) et du Saguenay-Lac-St-Jean (Bouchard et al., 1995; Heyer, 1995; Heyer et Tremblay, 1995).
La présente étude vise à estimer les effets de ce phénomène migratoire récent au Québec, dans une région particulièrement touchée, Lanaudière, située au nord-est de la région métropolitaine de Montréal (voir figure 6). À l’aide de l’approche généalogique, elle a pour but de caractériser la variabilité intra-régionale et les récentes transformations de son pool génique.
Population étudiée
Le peuplement d’origine européenne de Lanaudière a commencé sous le Régime français. Bien que trois seigneuries aient été concédées en 1640, ce n’est qu’en 1670 que des efforts sont déployés pour attirer des colons, et que neuf seigneuries additionnelles, situées sur la rive-nord du fleuve St-Laurent, sont octroyées. En 1736, toute la rive du fleuve est concédée (Morissonneau 1985; Tessier, 1987). Après la Conquête britannique de 1760, quelques cantons viendront s’ajouter dans les piedmonts et le plateau laurentien, où se sont notamment installés des Irlandais et des Écossais. Suite à leur déportation, de nombreuses familles acadiennes se sont établies dans quelques paroisses du sud de la région, telles que l’Assomption, l’Épiphanie et St-Jacques de l’Achigan (Hébert, 1994). La croissance démographique et l’essor de l’industrie forestière aux 18e et 19e siècles entraînent la fondation de nouvelles paroisses dans les territoires déjà colonisés. Afin de contrer l’exode rural de la fin du 19e siècle, le curé Brassard et l’abbé Provost incitent les colons à s’installer encore plus au nord, dans les « hauts », trois nouvelles paroisses sont ainsi fondées (Morissonneau, 1985).
Durant la deuxième moitié du 20e siècle, la population de Lanaudière a substantiellement augmenté dans un contexte où les taux de natalité ont chuté partout dans la province et où de nouveaux mouvements de population ont pris place (ISQ, 2008). En effet, entre 1951 et 1991, la population a triplé, passant de 105 821 à 335 372 habitants. Cette croissance a été plus importante dans les trois municipalités régionales de comté (MRC) bordant la région métropolitaine au sud de la région que dans les trois MRC du nord. Ce phénomène d’urbanisation en périphérie des grands centres a transformé le portrait du sud de Lanaudière. La population majoritairement rurale en 1951, se retrouve concentrée dans quelques agglomérations en 1991, comptant environ la moitié de la population (Asselin 1998, Rousseau, 2003). Cette expansion démographique est principalement expliquée par l’établissement de migrants en provenance des autres régions du Québec (Guillemette et St-Cyr, 1995; Rousseau, 2003). En accord avec ces changements démographiques, nous avons divisé la région de Lanaudière en deux sous-régions. La sous-région nord regroupe les MRC D’Autray, Joliette et Matawinie, et la sous-région sud, les MRC L’Assomption, Les Moulins et Montcalm).
Sources et structure des données
Nous avons construit quatre échantillons de cent individus de façon à représenter la répartition géographique de la population au sein de chaque sous-région, pour chacune des deux périodes étudiées, soit 1945-55 et 1985-95 (figures 1 et 2). Pour ce faire, nous avons utilisé la distribution municipale de la population tirée des recensements de 1951 et 1991. Ces individus, désignés comme points de départ des généalogies, ont été sélectionnés au hasard selon une méthode systématique parmi les mentions de mariages tirées de répertoires paroissiaux (SGL, 2008) et du fichier BALSAC-RETRO (Bouchard, 2007). Un seul individu a été sélectionné par couple, et nous avons alternativement choisi l’époux et l’épouse afin d’obtenir un nombre équivalent d’individus féminins et masculins. La distribution des sujets sélectionnés selon leurs lieux de mariage (figures 1 et 2) fait ressortir les municipalités les plus populeuses et la dispersion de la population sur le territoire de chacune des sous-régions, en 1945-55 et en 1985-95. Dans le but de faciliter la comparaison, nous avons créé des échantillons de même taille, bien que la répartition de l’ensemble de la population de Lanaudière dans les sous-régions nord et sud ne soit pas dans un rapport de un au cours des périodes étudiées, mais environ de 2 : un en 1951 et l’inverse en 1991.
Figure 1
Distribution géographique des sujets mariés en 1945-55

Figure 2
Distribution géographique des sujets mariés en 1985-95

Les généalogies ascendantes de ces individus ont été reconstruites à l’aide du fichier généalogique BALSAC-RETRO qui tire l’essentiel de ses données de deux fichiers sources, soit le fichier de population BALSAC et le Registre de population du Québec ancien (Bouchard, 2007; GRIG, 2008; PRDH, 2008). Les branches généalogiques ont été retracées aussi loin que les sources le permettent, c’est-à-dire jusqu’aux premiers arrivants sur le territoire québécois. La génération moyenne à laquelle les branches généalogiques s’interrompent est donnée par la mesure de la profondeur généalogique moyenne. Cette mesure considère le nombre de mentions d’ancêtres observées par rapport au nombre de mentions attendues à une génération donnée. Elle a été calculée de la façon proposée par Cazes et Cazes (1996).
Les généalogies des individus retenus dans nos échantillons sont complètes jusqu’à la deuxième génération, soit celle des grands-parents, afin de s’assurer que les données présentent un minimum de complétude. Les branches généalogiques ascendantes des individus mariés en 1945-55 atteignent en moyenne 9,3 générations, alors que celles des individus mariés en 1985-95 atteignent en moyenne 10,4 et 10,5 générations (tableau 1). En considérant un intervalle intergénérationnel moyen de trente ans (Tremblay et Vézina, 2000), ces valeurs indiquent que la majorité des généalogies remontent jusqu’aux fondateurs du 17e siècle. Le nombre d’ancêtres attendus dans une généalogie double à chaque génération. Il n’est donc pas surprenant d’observer que les mentions d’ancêtres dans les généalogies de la période plus récente soient plus du double qu’à la période précédente.
Tableau 1
Caractéristiques descriptives des corpus généalogiques

À cause de la structure arborescente des ascendances généalogiques et le nombre fini d’individus constituant une population, certains ancêtres apparaissent plus d’une fois dans les généalogies. Le nombre d’ancêtres distincts s’obtient alors en comptant chaque ancêtre une seule fois, indépendamment de son nombre d’apparitions (Vézina et al., 2005). Le rapport du nombre total de mentions d’ancêtres sur le nombre d’ancêtres distincts au sein de chacun des corpus généalogiques donne le nombre moyen d’occurrences des ancêtres et constitue un indice de l’homogénéité ancestrale des échantillons. En somme, les différentes caractéristiques descriptives des deux corpus généalogiques de Lanaudière de la période 1945-55 sont équivalentes (tableau 1). En plus d’attester de la comparabilité de ces corpus, l’équivalence des mesures descriptives représente un premier indicateur montrant que la structure généalogique des populations sous-régionales est similaire. En contraste, dans les corpus de la période 1985-95, les nombres de mentions d’ancêtres et d’ancêtres distincts sont plus élevés au sud qu’au nord de Lanaudière. De plus, les ancêtres apparaissent en moyenne moins souvent dans les généalogies du sud que dans celles du nord, indiquant un plus faible partage d’ancêtres et une plus grande diversité ancestrale dans cette sous-région.
Apparentement
Deux individus sont biologiquement apparentés s’ils ont un ou plusieurs ancêtres en commun. Du point de vue de la génétique, ceci signifie qu’ils ont une probabilité non nulle de partager un allèle (forme particulière d’un gène) hérité d’un ancêtre commun. Cette probabilité correspond au coefficient d’apparentement qui tient compte du nombre d’évènements de transmission de chacune des boucles généalogiques passant d’un individu à l’autre par un ancêtre commun, ainsi que du coefficient de consanguinité de cet ancêtre commun. Le coefficient d’apparentement est donc une mesure de distance génétique entre deux individus (Malécot, 1948).
Dans le but de saisir l’intensité des liens ancestraux unissant les sous-populations étudiées, le coefficient moyen d’apparentement a été calculé pour toutes les paires d’individus possibles à l’intérieur d’un groupe donné (intragroupe), mais aussi entre deux groupes donnés (intergroupe) et pour chaque profondeur générationnelle. Ainsi, pour deux corpus de cent sujets, la moyenne des coefficients d’apparentement des 4950 paires d’individus a été calculée à l’intérieur de chaque groupe, de même que celle des 10 000 coefficients possibles entre les individus des deux groupes. Les comparaisons statistiques entre les coefficients moyens d’apparentement ont été réalisées à l’aide d’un test de permutation (Lavoie et al., 2005). La statistique du test a été obtenue grâce à 5000 répétitions afin d’obtenir une précision suffisante au seuil de signification α = 0,05. L’apparentement observé sur un nombre restreint de générations (jusqu’à 5) est généralement désigné comme un apparentement proche, alors que pour un nombre plus élevé on parle d’apparentement éloigné.
Les coefficients moyens d’apparentement observés au sein de chaque échantillon et entre les paires de groupes sont représentés dans les figures 3A et 3B. Les coefficients sont nuls lorsque seule la génération des parents est considérée puisque nous avons intentionnellement exclu les paires de frères et soeurs de nos échantillons. Les courbes d’apparentement croissent jusqu’à la 11e ou la 12e génération où elles atteignent un plateau, indiquant que peu de nouveaux liens généalogiques ont pu être ensuite identifiés. L’essentiel de l’augmentation des coefficients se situe après la 6e génération, ce qui souligne l’importance de l’apparentement éloigné dans les sous-populations lanaudoises, tel qu’il a été précédemment établi pour l’ensemble des populations régionales québécoises (Tremblay et al., 2001; Vézina et al., 2005; Vézina et al., 2006).
Figure 3
Coefficients d’apparentement moyens (x 10000) intragroupe (A) et intergroupe (B) par génération
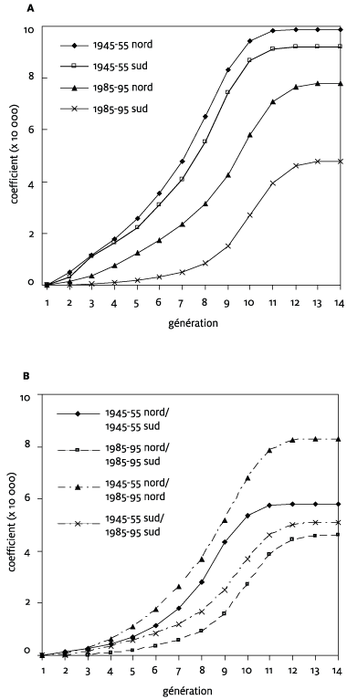
Les échantillons nord et sud de la période 1945-55 montrent un apparentement intragroupe moyen similaire à toutes les profondeurs générationnelles (p > 0,1) et affichent les coefficients les plus élevés qui atteignent les maximums respectifs de 9,20 et 9,86 pour 10000 (figure 3A). Ces valeurs correspondent à l’apparentement entre cousins du quatrième degré. Cet apparentement est équivalent à celui observé par Vézina et al. (2005) pour un échantillon quatre-vingt-dix individus lanaudois mariés entre 1935 et 1974, et qui se situait parmi les apparentements les plus faibles observés dans les vingt-six régions étudiées. D’une période à l’autre, les coefficients d’apparentement sont significativement réduits dans les deux sous-régions (p < 0,05). Les niveaux d’apparentement intergroupe sont aussi globalement plus faibles que ceux intragroupe observés pour la période 1945-55. Les individus d’une sous-région donnée mariés en 1945-55 sont donc plus apparentés entre eux qu’ils ne le sont avec les individus de l’autre sous-région, ou de l’autre période. Ceci signifie qu’il existe une certaine différenciation génétique entre ces deux sous-régions, qui pourrait résulter de leur contexte historique de peuplement. Cette différence réside surtout dans les générations proches où les valeurs sont presque nulles entre les sous-régions.
Pour la période 1985-95, les sujets du sud de Lanaudière sont nettement moins apparentés entre eux que ceux du nord. De plus, les sujets du sud ne sont pas plus apparentés entre eux qu’avec les sujets du nord. Ces mesures d’apparentement représentent une autre démonstration de la plus grande hétérogénéité génétique du sud de Lanaudière. L’apparentement intragroupe au nord de Lanaudière augmente progressivement à partir de la deuxième génération, et atteint une valeur maximale de 7,78 pour 10 000. En comparaison, les liens qui unissent les sujets du sud sont moins nombreux à passer par des ancêtres ayant vécu aux 19e et 20e siècles, tandis qu’une grande proportion de ces liens correspond à la période du Régime français (8e à 12e génération). Ainsi, la vaste majorité des lignées ascendantes du sud, et une bonne proportion de celles du nord, ne se sont pas métissées entre elles depuis la Conquête. L’évolution de ces lignées s’est donc déroulée dans des contextes relativement séparés durant quelques générations suivant l’expansion de la population canadienne française dans les différentes régions du Québec, et leur entrée dans Lanaudière est récente.
Consanguinité
Un individu est dit issu d’une union consanguine lorsque ses parents sont liés dans leurs ascendances par un ou plusieurs ancêtres communs (Wright, 1922). Le coefficient de consanguinité F correspond alors à la probabilité qu’un individu ait hérité pour un gène donné de deux allèles identiques par ascendance, l’un provenant de sa mère, et l’autre de son père. Afin d’étudier ce phénomène au sein des sous-populations lanaudoises, nous avons évalué le coefficient moyen de consanguinité, qui est la moyenne des coefficients de consanguinité des individus d’un groupe donné. Les valeurs obtenues ont été comparées à l’aide d’un test de Student. L’analyse des coefficients par génération permet de distinguer la consanguinité proche, qui nous informe des modalités du choix du conjoint, de la consanguinité éloignée qui, au même titre que l’apparentement, nous renseigne sur la structure ancestrale de la population. La principale conséquence de la consanguinité au sein d’une population est une augmentation de la proportion des génotypes homozygotes aux dépens des hétérozygotes (Hartl, 1994). Comme la diversité génétique peut être qualifiée par la proportion d’hétérozygotie, un plus faible coefficient de consanguinité correspond à une plus grande diversité.
Figure 4
Coefficient moyen de consanguinité (x 10000) (A) et proportion d’individus (%) issus d’une union consanguine (B) par génération

Les coefficients moyens de consanguinité (figure 4A) augmentent jusqu’à la onzième génération où ils atteignent un plateau. À ce niveau de génération, 99 % ou plus des individus sont issus d’une union consanguine (figure 4B). Ce résultat souligne l’importance de la consanguinité de type éloigné dans les corpus de Lanaudière, comparativement à la consanguinité proche, qui ne concerne qu’un faible nombre d’individus. Néanmoins, ce constat n’est pas unique à Lanaudière, il concerne toutes les populations régionales du Québec (Vézina et al., 2005).
Les coefficients de consanguinité sont globalement plus élevés que les coefficients d’apparentement au sein des corpus de Lanaudière (figure 3A). Autrement dit, les conjoints sont plus apparentés entre eux que la population générale. On qualifie ce phénomène d’endogamie et il s’explique par le choix préférentiel des conjoints à l’intérieur d’une sous-population donnée. Du point de vue de la génétique des populations, lorsque les individus choisissent leurs conjoints en fonction de la proximité géographique, ceci entraîne une réduction de la variabilité génétique au sein des populations et un phénomène de différentiation génétique entre les populations.
Le corpus du nord de Lanaudière en 1945-55 montre des niveaux de consanguinité nettement plus élevés que les autres corpus à toutes les profondeurs générationnelles (valeur maximale = 74,5). Cette différence s’explique principalement par des boucles de consanguinité proche chez quelques individus aux troisième et quatrième générations (figure 4B). Cette endogamie plus importante au nord de Lanaudière peut être issue de la plus grande distance séparant les villages sur ce territoire, par rapport au sud, y créant un marché nuptial plus réduit, ou de pratiques locales de choix du conjoint. Après la quatrième génération, les coefficients des sous-régions lanaudoises augmentent proportionnellement (p > 0,05), et le nombre de sujets ayant un coefficient de consanguinité non-nul est alors comparable.
De la période plus ancienne à la plus récente, on observe une réduction significative des niveaux de consanguinité proche et éloignée au nord de Lanaudière, et entre les huitième et dixième générations au sud (p > 0,05). Ce phénomène va de pair avec le plus faible nombre d’individus issus d’une union consanguine jusqu’à la dixième génération. De plus, le rapport entre les sous-régions change. En effet, le nord de Lanaudière montre des niveaux de consanguinité proche nettement plus forts que le sud de Lanaudière entre 1945-55, alors que les coefficients de la période 1985-95 ne sont pas significativement différents. Somme toute, ces résultats suggèrent une plus grande hétérogénéité génétique dans les sous-régions lanaudoises en 1985-95.
Contribution génétique des fondateurs
La variabilité intrarégionale de Lanaudière a été davantage caractérisée par l’analyse de la contribution génétique des fondateurs immigrants et régionaux. À chaque génération, un parent transmet la moitié de son génome à chacun de ses enfants. Ainsi, en remontant les générations, il est possible de calculer la part du génome d’un individu ou d’un groupe d’individus provenant d’un fondateur donné. La constitution génétique d’une population peut alors être spécifiée par l’arrangement particulier des contributions génétiques de ses fondateurs (Roberts, 1968).
Nous avons analysé la contribution génétique des fondateurs immigrants et régionaux selon leur origine géographique et leur période de mariage. Ce dernier paramètre nous permet d’estimer la période d’implantation des fondateurs sur le territoire considéré. Lorsqu’un fondateur a été marié plus d’une fois, nous avons retenu l’année moyenne de ses mariages. Dans les quelques cas où la date de mariage d’un fondateur était indéterminée, nous avons défini son année de mariage comme l’année moyenne de mariage de ses enfants moins trente ans, qui équivaut à la durée moyenne d’une génération dans la population canadienne française et dans les populations occidentales (Tremblay et Vézina, 2000; Fenner, 2005).
Fondateurs immigrants
Les fondateurs immigrants sont définis comme les premiers ancêtres immigrants sur le territoire québécois identifiés dans l’ensemble des lignées ascendantes. Leurs origines géographiques ont été tirées directement des sources disponibles sur leurs lieux de naissance, de mariage ou d’émigration ou indirectement désignées à partir de celles de leurs parents. Pour 1945-55, nous avons retracé 3151 fondateurs immigrants dans la sous-région nord et 3211 au sud. Parmi ceux-ci, 2602 sont communs aux deux sous-régions et les fondateurs spécifiques restants se retrouvent à parts égales au nord et au sud. Pour 1985-95, 3947 fondateurs immigrants ont été identifiés au nord de Lanaudière et 4354 au sud. Pour cette période, 3492 se retrouvent dans les deux sous-régions, mais on retrouve un nombre plus élevé de fondateurs spécifiques au sud (862) qu’au nord (455). Le plus grand nombre de fondateurs immigrants dans les corpus de 1985-95 s’explique par la génération supplémentaire atteinte dans ces généalogies.
Les fondateurs immigrants spécifiques à chaque corpus se sont majoritairement mariés au cours du 18e siècle (figure 5A). La contribution de ces fondateurs est semblable dans les corpus de la période 1945-55 (nord 2,3 % et sud 3 %), tandis qu’elle est plus marquée au sud de Lanaudière en 1985-95 (nord 3,2 % et sud 6,4 %) (figure 5B). Dans l’ensemble, les fondateurs immigrants ayant le plus contribué aux pools géniques sous-régionaux sont ceux mariés au 17e siècle. Dans chacun des quatre échantillons, un peu plus de 80 % des pools géniques provient de ces fondateurs. Au second rang viennent les fondateurs mariés au 18e siècle (~ 15 %). Enfin, la contribution des fondateurs mariés après 1800 représente une faible part des pools géniques des sujets de Lanaudière. La période initiale du peuplement a donc été déterminante dans la constitution génétique de la population contemporaine de Lanaudière.
Figure 5
Distribution (A) et contribution génétique (%) (B) des fondateurs immigrants selon leur période de mariage (la part hachurée représente les fondateurs spécifiques)

La contribution des fondateurs immigrants aux pools géniques de 1985-95 selon leur origine est très similaire à celle observée quarante ans plus tôt (tableau 2). Elle reflète la composition de l’immigration fondatrice des 17e et 18e siècles, où les fondateurs venus directement de France sont de loin les plus importants (~ 90 %) (Charbonneau et al., 2000). Au second rang viennent les fondateurs d’origine acadienne (~ 5 %) et enfin, ceux d’autres pays d’Europe et d’ailleurs en Amérique (~ 5 %). Cette distribution est conforme à ce qui a été décrit pour Lanaudière dans l’étude préliminaire de Tremblay et al. (2003) ainsi que pour l’ensemble du Québec par Vézina et al. (2006). Ces résultats suggèrent une empreinte de l’effet fondateur initial qui perdure dans Lanaudière, malgré les récents changements démographiques.
Tableau 2
Distribution et contribution génétique totale (%) des fondateurs immigrants selon leur origine
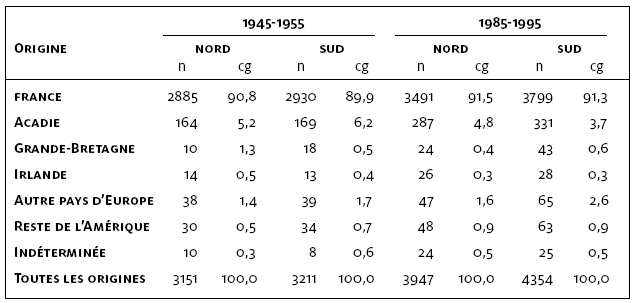
Les différences observées entre les sous-régions nord et sud se retrouvent dans le nombre et la contribution génétique des fondateurs qui proviennent d’un autre pays que la France, mais elles ont une faible incidence sur l’ensemble des pools géniques sous-régionaux étant donné leurs modestes valeurs. Dans les corpus de la période plus ancienne, 164 et 169 fondateurs immigrants acadiens sont dénombrés au nord et au sud de Lanaudière, où respectivement 5,2 % et 6,2 % des bassins génétiques leur sont attribuables. Ces observations sont équivalentes à celles de Bergeron et al. (2008) qui ont étudié la contribution des Acadiens au sein de vingt-six régions du Québec, chacune représentée par quatre-vingt-dix individus mariés entre 1935 et 1974.
Fondateurs régionaux
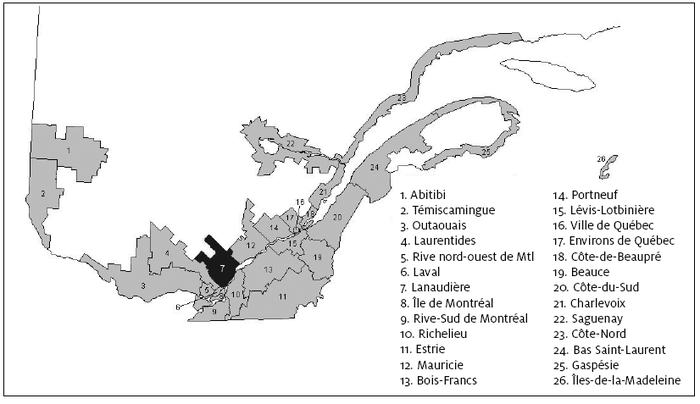
Les fondateurs régionaux sont définis comme les premiers ancêtres à s’être mariés dans Lanaudière, leurs parents s’étant mariés dans une autre région du Québec ou à l’extérieur de la province. Le lieu d’origine d’un fondateur régional québécois a été désigné comme étant la région de mariage de ses parents selon les divisions territoriales utilisées par le GRIG (figure 6). La caractérisation des fondateurs régionaux révèle des informations intéressantes sur les origines et la structuration des populations régionales. Des travaux ont déjà été effectués à ce sujet pour les régions de l’Abitibi et du Témiscamingue (Bilodeau, 2002) et les sous-régions du Bas-Saguenay, du Haut-Saguenay et du Lac-St-Jean (Lavoie et al., 2005).
Figure 6
Découpage géographique du territoire du Québec en 26 régions
Au total, 2475 et 2889 fondateurs régionaux ont été identifiés dans les généalogies de 1945-55 et 1985-95, respectivement, dont 1363 spécifiques à un seul corpus, 891 communs à deux corpus, 615 communs à trois corpus et 585 communs aux quatre corpus. En comparaison avec les fondateurs immigrants, on observe un moindre nombre absolu de fondateurs régionaux et une plus grande proportion de fondateurs spécifiques au détriment des fondateurs communs à plusieurs corpus. Les fondateurs régionaux mariés avant 1700 se retrouvent en faible nombre dans les corpus de Lanaudière (au total : 56) (figure 7A). Les plus anciens observés se sont mariés en 1672. Plus de 80 % des fondateurs régionaux se sont mariés entre 1700 et 1800, et moins de 15 % après 1800. La forte concentration des mariages fondateurs au 18e siècle coïncide avec la période initiale de peuplement de Lanaudière, qui a réellement commencé près d’un siècle après l’arrivée des premiers immigrants européens en Nouvelle-France (Morissonneau, 1985). Bon nombre de fondateurs immigrants sont donc plus anciens (en terme de générations les séparant des sujets) que les fondateurs régionaux de Lanaudière.
Figure 7
Distribution (A) et contribution génétique (%) (B) des fondateurs régionaux selon leur période de mariage (la part hachurée désigne les fondateurs spécifiques)

La contribution génétique des fondateurs régionaux selon leur période de mariage est équivalente dans les deux corpus de la période 1945-55 (figure 7B). Les fondateurs régionaux mariés au 18e siècle sont les plus importants en nombre et contribuent pour 70 % des pools géniques sous-régionaux de 1945-55. En ordre décroissant, suivent les fondateurs mariés au 19e siècle (~ 17 %), après 1900 (~ 10 %), et entre 1672 et 1700 (~ 4 %). Des contributions similaires valent pour la période suivante au nord de Lanaudière, mais elles diffèrent au sud. En effet, dans la sous-région sud, en 1985-95, la contribution des fondateurs régionaux mariés après 1900 (52 %) surpasse celle des fondateurs du 18e siècle (36 %). Les fondateurs récents sont aussi plus nombreux dans ce corpus que dans les trois autres, 92 % d’entre eux sont les parents d’un sujet. Ceux-ci ont la plus forte contribution possible (0,5), mais ne sont liés qu’à un seul sujet, ce qui leur confère un faible apport individuel. La moitié du pool génique de la sous-région sud est donc attribuable à des fondateurs établis dans Lanaudière au cours du 20e siècle.
La contribution génétique des fondateurs régionaux groupés selon leurs origines présente une variabilité considérable d’un corpus à l’autre (tableau 3). Les fondateurs régionaux proviennent soit d’une autre région du Québec, soit de l’extérieur du Québec. Comme on ne peut déterminer la région au Québec où les contrats de mariage ont été alloués, nous les avons regroupés dans une catégorie à part. Les fondateurs régionaux dont le lieu d’origine est indéterminé, contribuent pour une part non négligeable aux pools géniques sous-régionaux. Une faible proportion de fondateurs immigrants (~ 10 %) s’est directement établie dans Lanaudière. La majorité d’entre eux s’est d’abord installée ailleurs en Nouvelle-France, puis leurs descendants québécois ont migré vers Lanaudière. Étonnamment, les fondateurs régionaux français et acadiens sont d’égale importance démographique et génétique dans les corpus de Lanaudière. Les fondateurs régionaux originaires d’un autre pays sont peu nombreux et leur contribution génétique ne dépasse pas 1,5 %. Ceci pourrait expliquer, du moins en partie, l’influence majeure des Acadiens sur la culture populaire de Lanaudière (Morissonneau, 1985). Par ailleurs, notons que bien que les premières familles acadiennes de Lanaudière se soient établies principalement au sud de Lanaudière (Morissonneau, 1985; Hébert, 1994), elles ont également contribué aux pools géniques des sous-régions nord et sud.
Tableau 3
Distribution et contribution génétique totale (%) des fondateurs régionaux selon leur origine
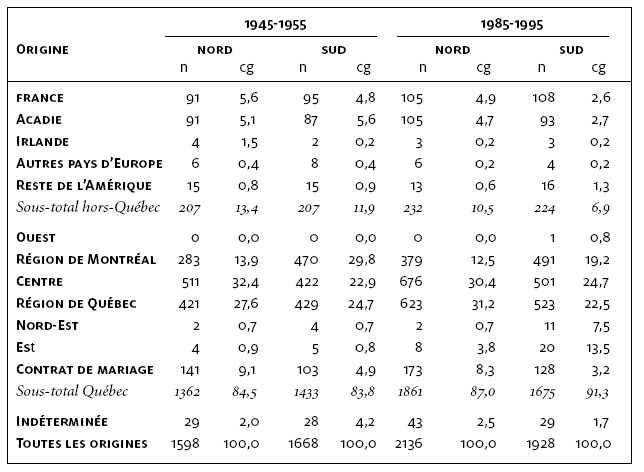
Note : Les 26 régions du Québec (figure 6) ont été regroupées en 6 ensembles régionaux afin de faciliter l’analyse. Ouest : 1 à 4; Région de Montréal : 5, 6, 8 et 9; Centre : 10 à 13; Région de Québec : 14 à 20; Nord-Est : 21 à 23; Est : 24 à 26. Les régions du Québec où les contrats de mariage ont été établis n’ont pu être déterminées.
L’analyse des fondateurs régionaux de Lanaudière met à nouveau en évidence l’importance du peuplement initial de la région sur la structure de son pool génique en 1945-55. La distribution des fondateurs régionaux de Lanaudière selon leur région d‘origine reproduit la répartition géographique de la population durant la période initiale du peuplement de ce territoire, laquelle était organisée autour de trois principaux ports d’entrée des immigrants – Montréal, Québec et Trois-Rivières — (Charbonneau et al., 1987). Pour la période 1945-55, environ 75 % des pools géniques sous-régionaux proviennent des fondateurs régionaux issus de ces trois régions (centre du Québec, région de Montréal et de Québec). Néanmoins, leur contribution aux sous-régions varie en fonction de la proximité géographique. Les fondateurs en provenance de la région de Montréal contribuent plus au pool génique du sud de Lanaudière, tandis qu’au nord, une plus grande part du pool génique provient des fondateurs originaires du Centre du Québec et de la région de Québec. Moins de 2 % de ces pools géniques sont issus des fondateurs régionaux de l’Est et du Nord-Est du Québec.
Les variations de la part du pool génique attribuable aux fondateurs régionaux d’une origine et d’une période de mariage données reflètent l’évolution des comportements migratoires interrégionaux entre les deux périodes. À cet égard, la structuration du pool génique du nord de Lanaudière en 1985-95 est semblable à celle observée quarante ans plus tôt dans la même sous-région. En effet, 61,6 % du pool génique est attribuable aux fondateurs régionaux provenant du Centre du Québec et de la région de Québec, 12,5 % provient de fondateurs de la région de Montréal. Cependant, une plus grande part du pool génique est attribuable aux fondateurs originaires de l’Est du Québec (3,8 %). Les changements dans la composition du pool génique de la sous-région sud sont davantage accentués. Bien que la proportion de fondateurs régionaux provenant de l’extérieur du Québec et de la région de Montréal change peu d’une période à l’autre, leur contribution génétique est réduite (respectivement de 6,9 % et 19,2 %), au profit des fondateurs régionaux originaires de l’Est (13,5 %) et du Nord-Est québécois (7,5 %). Ces derniers se sont manifestement installés au sud de Lanaudière récemment en suivant le mouvement migratoire vers la périphérie de la région métropolitaine.
Discussion
La population de Lanaudière, qui était dispersée en 1951, a énormément crû au cours des dernières décennies grâce aux migrations interrégionales et s’est progressivement agglomérée au sud dans quelques grands centres urbains en périphérie de la région métropolitaine (Asselin, 1998; Rousseau, 2003). En nous appuyant sur des observations réalisées au sein d’autres populations urbaines (Helgason et al., 2005; Ashrafian-Bonab et al., 2007), nous avons testé, premièrement, l’hypothèse que ces migrations se traduisent par une augmentation de la diversité et une réduction de la stratification génétique (différentiation entre les sous-régions) et, deuxièmement, que leurs effets sont plus marqués dans le sud que dans le nord. En effet, plusieurs mesures démogénétiques indiquent une augmentation de l’hétérogénéité du pool génique du sud de Lanaudière, en contraste avec celui du nord pour lequel l’intensité des transformations est sensiblement moindre. De plus, à l’opposé des individus mariés en 1945-55, ceux mariés en 1985-95 dans la sous-région sud ne sont pas plus apparentés entre eux qu’avec les autres groupes. Autrement dit, ce pool génique est moins différencié que celui du nord et que ceux de 1945-55. En 1985-95, la moitié du bassin génétique du sud de Lanaudière est issu de fondateurs régionaux spécifiques et récents qui sont principalement originaires d’autres régions du Québec. Bien que des analyses moléculaires soient nécessaires pour estimer la variabilité génétique introduite par ces nouveaux fondateurs régionaux, ces résultats doivent être pris en compte dans les stratégies d’échantillonnage. Par exemple, une étude de cartographie génétique portant sur une population urbaine devrait être accompagnée d’une collecte de données démographiques sur les origines géographiques des parents et grands-parents, comme suggéré par Ashrafian-Bonad et al. (2007).
Malgré les transformations et les mouvements de population récents, un résultat intéressant de cette étude est que l’importance des fondateurs immigrants de l’époque de la Nouvelle-France perdure au cours du 20e siècle dans Lanaudière malgré les transformations et les mouvements de population récents. Ceci souligne donc que les nouveaux arrivants dans Lanaudière en provenance des autres régions du Québec sont bel et bien les descendants d’un même pool original de fondateurs et que leur importance relative – en terme de contribution génétique – était déterminée avant l’expansion de la population dans les différentes régions. Néanmoins, rappelons que nos données proviennent essentiellement des registres catholiques et qu’elles ne permettent d’estimer l’apport des immigrants plus récents que dans la mesure où ils se sont intégrés à la population canadienne française par le biais d’un mariage. Par ailleurs, notre étude souligne aussi l’importance de la structure géographique d’une population au moment de la colonisation d’un nouveau territoire. Dans le cas de Lanaudière, l’organisation en trois pôles majeurs de la population du Québec ancien (Montréal, Québec et Trois-Rivières) est reflétée par l’importance des fondateurs régionaux en provenance de ces régions. La structure tripartite du pool génique québécois a déjà été relevée dans des études précédentes (Gagnon et Heyer, 2001; Tremblay et al., 2001; Vézina et al., 2005; Bergeron et al., sous presse).
On observe donc au cours du 20e siècle une augmentation de la diversité des pools géniques lanaudois, correspondant à une réduction des coefficients d’apparentement et de consanguinité, une réduction de la structuration de la population de la sous-région sud et une modification des trajectoires migratoires, telles qu’indiquées par les origines et périodes de mariage des fondateurs régionaux. Les migrations vers la périphérie des zones urbaines attirent non seulement les individus des régions adjacentes, mais aussi ceux de régions plus éloignées de l’est et du nord-est de la province. Bien que nous n’ayons examiné que la population canadienne française de Lanaudière, il y a fort à parier que ces tendances se retrouvent aussi dans les régions ceinturant l’île de Montréal qui ont subi une expansion démographique similaire.
Enfin, la présente étude a également mis en lumière l’importance de la composante géographique dans la structure génétique d’une population contemporaine. Pour y faire suite, il serait intéressant d’évaluer l’importance et les conséquences de l’expansion de la population québécoise dans les différentes régions québécoises, afin d’expliquer, notamment, la régionalisation de l’effet fondateur initial.
Appendices
Remerciements
Les auteurs remercient les assistants de recherche du GRIG et du Projet BALSAC pour leur assistance technique ainsi que le Laboratoire de géographie historique de l’Université Laval pour la production des cartes. Cette étude a reçu l’appui financier du Réseau de médecine génétique appliquée du Québec (RMGA-FRSQ).
Bibliographie
- Ashrafian-Bonab, M., L. J. Lawson Handley et F. Balloux. 2007. « Is urbanization scrambling the genetic structure of human populations? A case study », Heredity, 98, 3 : 151-156.
- Asselin, R. 1998. Lanaudière : la région administrative, les MRC et les principales municipalités : Recensements de la population : 1996-1991-1986 : Données comparatives et faits saillants. Cahier 1 : Population et logement. Direction de la géostatistique et de l’information, Bureau de la Statistique du Québec, 128 p.
- Bergeron, J., H. Vézina, L. Houde et M. Tremblay. 2008. « La contribution des Acadiens au peuplement des régions du Québec », Cahiers québécois de démographie, 37, 1 : 181-204.
- Bilodeau, M. 2002. Caractéristiques démogénétiques des populations de l’Abitibi et du Témiscamingue. Mémoire de maîtrise, Université Laval/Université du Québec à Chicoutimi, 81 p.
- Bouchard, G. 2007. Projet BALSAC – Rapport annuel 2006-2007. Chicoutimi (Québec), Canada, 58 p.
- Bouchard, G., H. Charbonneau, B. Desjardins, E. Heyer, et M. Tremblay. 1995. « Mobilité géographique et stratification du pool génique canadien-français sous le Régime français », dans Y. Landry, J. Dickinson, S. Pasleau et C. Desama, éd. Les chemins de la migration en Belgique et au Québec xviie – xxe siècles. Beauport, Publications MNH : 51-59.
- Campbell, H., A. D. Carothers, I. Rudan, C. Hayward, Z. Biloglav, L. Barac, M. Pericic, B. Janicijevic, N. Smolej-Narancic, O. Polasek, I. Kolcic, J. L. Weber, N. D. Hastie, P. Rudan et A. F. Wright. 2007. « Effects of genome-wide heterozygosity on a range of biomedically relevant human quantitative traits », Human Molecular Genetics, 16, 2 : 233-241.
- Cavalli-Sforza, L. L., A. C. Wilson, C. R. Cantor, R. M. Cook-Deegan et M. C. King. 1991. « Call for a worldwide survey of human genetic diversity : a vanishing opportunity for the Human Genome Project », Genomics, 11, 2 : 490-491.
- Cazes, M.-H., et P. Cazes. 1996. « Comment mesurer la profondeur généalogique d’une ascendance? », Population, 51, 1 : 117-140.
- Charbonneau, H., B. Desjardins, A. Guillemette, Y. LandrY, J. Légaré et F. Nault. 1987. Naissance d’une population. Les Français établis au Canada au xviie siècle. Paris/Montréal, Presses universitaires de France/Presses de l’Université de Montréal, 232 p.
- Charbonneau, H., B. Desjardins, J. Légaré et H. Denis. 2000. « The population of the St-Lawrence Valley, 1608-1760 », dans M. R. Haines et R. H. Steckel, éd. A population history of North America. Cambridge, Cambridge University Press : 99-142.
- Clark, D. 2003. Urban World, Global City (2nd edition). London, Routledge, 235 p.
- Fenner, J. N. 2005. « Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies », American Journal of Physical Anthropology, 128, 2 : 415-423.
- Gagnon, A., et E. Heyer. 2001. « Fragmentation of the Quebec population genetic pool (Canada) : evidence from the genetic contribution of founders per region in the 17th and 18th centuries », American Journal of Physical Anthropology, 114, 1 : 30-41.
- Grig. Groupe de recherche interdisciplinaire en démographie et épidémiologie génétique.http://www.uqac.ca/grig/ (page consultée le 01/07/2008).
- Guillemette, A., et J.-P. St-Cyr. 1995. Mosaïque lanaudoise. Profil démographique, socio-économique et sanitaire de la population lanaudoise. Joliette, Direction de la santé publique, Régie régionale de la santé et des services sociaux de Lanaudière, 130 p.
- Hartl, D. 1994. Génétique des populations. Paris, Flammarion, 305 p.
- Hébert, P.-M. 1994. Les Acadiens du Québec. Montréal, Éditions de l’Écho, 480 p.
- Helgason, A., B. Hrafnkelsson, J. R. GulcheR, R. Ward et K. Stefansson. 2003. « A Populationwide Coalescent Analysis of Icelandic Matrilineal and Patrilineal Genealogies : Evidence for a Faster Evolutionary Rate of mtDNA Lineages than Y Chromosomes », American Journal of Human Genetics, 72, 6 : 1370-1388.
- Helgason, A., B. Yngvadottir, B. Hrafnkelsson, J. Gulcher et K. Stefansson. 2005. « An Icelandic example of the impact of population structure on association studies », Nature Genetics, 37, 1 : 90-95.
- Heyer, E. 1995. « Genetic consequences of differential demographic behaviour in the Saguenay region, Quebec », American Journal of Physical Anthropology, 98 : 1-11.
- Heyer, E., et M.-H. Cazes 1999. « Les “enfants utiles” : une mesure démographique pour la génétique des populations », Population 54, 4-5 : 677-691.
- Heyer, E., et M. Tremblay. 1995. « Variability of the genetic contribution of Quebec population founders associated to some deleterious genes », American Journal of Human Genetics, 56 : 970-8.
- ISQ. Institut de la statistique du Québec.http://www.stat.gouv.qc.ca/ (page consultée le 01/07/2008).
- Lavoie, E. M., M. Tremblay, L. Houde et H. Vézina. 2005. « Demogenetic study of three populations within a region with strong founder effects », Community Genetics, 8, 3 : 152-160.
- Malécot, G. 1948. Les mathématiques de l’hérédité. Paris, Masson, 60 p.
- Morissonneau, C. (Dir.). 1985. Guide de Lanaudière : Culture, histoire et tourisme. Joliette, 327 p.
- PRDH. Programme de recherche en démographie historique de l’Université de Montréal.http://www.prdh.umontreal.ca (page consultée le 01/07/2008).
- Roberts, D. F. 1968. « Genetic effects of population size reduction », Nature, 220, 172 : 1084-1088.
- Rousseau, N. 2003. « Socio-démographie de la région de Lanaudière (Québec) : recension des écrits et portrait statistique », Rapport de recherche présenté au GRIG, 242 p.
- Rudan, I., A. D. Carothers, O. Polasek, C. Hayward, V. Vitart, Z. Biloglav, I. Kolcic, L. Zgaga, D. Ivankovic, A. Vorko-Jovic, J. F. Wilson, J. L. Weber, N. Hastie, A. Wright et H. Campbell. 2008. « Quantifying the increase in average human heterozygosity due to urbanisation », European Journal of Human Genetics, Mar 5.
- SGL. Société de généalogie de Lanaudière.http://www.sgl.lanaudiere.net (page consultée le 01/07/2008).
- Tessier, D. 1987. Bibliographie de Lanaudière. Québec, Institut québécois de recherche sur la culture, 270 p.
- Tremblay, M., B. Desjardins, L. Houde et H. Vézina. 2003. Dimension régionale de l’effet fondateur québécois : le cas de Lanaudière. Rimouski, 71e Congrès de l’ACFAS, Colloque de l’ADQ, 19-23 mai.
- Tremblay, M., M. Jomphe et H. Vézina. 2001. « Comparaison de structures patronymiques et génétiques dans la population québécoise », dans G. Brunet, P. Darlu et G. Zei, dir. Le patronyme : histoire, anthropologie, société. Paris, Éditions du CNRS : 367-389.
- Tremblay, M., et H. Vézina. 2000. « New estimates of intergenerational time intervals for the calculation of age and origins of mutations », American Journal of Human Genetics, 66, 2 : 651-658.
- Vézina, H., M. Tremblay, B. Desjardins et L. Houde. 2005. « Origines et contributions génétiques des fondatrices et des fondateurs de la population québécoise », Cahiers québécois de démographie, 34, 2 : 235-258.
- Vézina, H., M. Tremblay et L. Houde. 2005. « Mesures de l’apparentement biologique au Saguenay-Lac-St-Jean (Québec, Canada) à partir de reconstitutions généalogiques », Annales de démographie historique, 2 : 67-84.
- Vitart, V., A. D. Carothers, C. Hayward, P. Teague, N. D. Hastie, H. Campbell et A. F. Wright. 2005. « Increased level of linkage disequilibrium in rural compared with urban communities : a factor to consider in association-study design », American Journal of Human Genetics, 76, 5 : 763-772.
- Wright, S. 1922. « Coefficient of inbreeding and relationship », American Naturalist, 56 : 330-338.
 10.7202/029644ar
10.7202/029644arList of figures
Figure 1
Distribution géographique des sujets mariés en 1945-55
Figure 2
Distribution géographique des sujets mariés en 1985-95
Figure 3
Coefficients d’apparentement moyens (x 10000) intragroupe (A) et intergroupe (B) par génération
Figure 4
Coefficient moyen de consanguinité (x 10000) (A) et proportion d’individus (%) issus d’une union consanguine (B) par génération
Figure 5
Distribution (A) et contribution génétique (%) (B) des fondateurs immigrants selon leur période de mariage (la part hachurée représente les fondateurs spécifiques)
Figure 6
Découpage géographique du territoire du Québec en 26 régions
Figure 7
Distribution (A) et contribution génétique (%) (B) des fondateurs régionaux selon leur période de mariage (la part hachurée désigne les fondateurs spécifiques)
List of tables
Tableau 1
Caractéristiques descriptives des corpus généalogiques
Tableau 2
Distribution et contribution génétique totale (%) des fondateurs immigrants selon leur origine
Tableau 3
Distribution et contribution génétique totale (%) des fondateurs régionaux selon leur origine
Note : Les 26 régions du Québec (figure 6) ont été regroupées en 6 ensembles régionaux afin de faciliter l’analyse. Ouest : 1 à 4; Région de Montréal : 5, 6, 8 et 9; Centre : 10 à 13; Région de Québec : 14 à 20; Nord-Est : 21 à 23; Est : 24 à 26. Les régions du Québec où les contrats de mariage ont été établis n’ont pu être déterminées.