Résumés
Résumé
L’objectif de cette étude consistait à déterminer les effets létaux et sublétaux de quatre formulations utilisées communément en cultures maraîchères de plein champ sur l’auxiliaire Episyrphus balteatus [Diptera : Syrphidae]. La toxicité aiguë et les effets chroniques de ces produits à la dose recommandée et à doses réduites furent évalués en conditions de laboratoire en plaçant les insectes directement en contact avec des organes végétaux imprégnés de pesticide. Les insecticides Karate® et Pirimor® se sont révélés très toxiques, même à faible dose. Une certaine sélectivité n’apparaît qu’à 2,5 % de la dose recommandée. Les fongicides Ronilan®et Quintalic®qui n’ont pas montré une toxicité aiguë aussi drastique ont néanmoins engendré des effets néfastes à plus long terme sur les paramètres de développement et de reproduction.
Abstract
The aim of this work was to assess both lethal and sub-lethal effects of four pesticides commonly used in field vegetables in south Belgium (Wallonia) on Episyrphus balteatus [Diptera: Syrphidae]. Acute toxicity and long term effects of the pesticides at the recommended and reduced doses were assessed in laboratory conditions by exposing the insects on plant leaves previously treated with the considered pesticide. Both tested insecticides, namely Karate® and Pirimor®, were found to be very harmful for the beneficial hoverfly even at reduced doses. A kind of selectivity only appeared from 2.5% of the recommended dose. The two fungicides, namely Ronilan® and Quintalic®, which showed a moderate acute toxicity had induced negative effects on long-term parameters, on both development and reproduction.
Corps de l’article
Introduction
Dans le cadre de la diversification des productions végétales, on observe en Belgique une tendance à l’accroissement des superficies consacrées aux cultures légumières. Ces produits horticoles doivent répondre à des normes de qualité, notamment en ce qui concerne les limites maximales de résidus de pesticides. Les méthodes alternatives de lutte, dont l’utilisation des auxiliaires, constituent un moyen de contrôle complémentaire qui permet de réduire le nombre d’interventions chimiques et les résidus de pesticides dans les denrées alimentaires.
Lors de la mise en oeuvre d’un programme de lutte intégrée basé sur l’utilisation combinée d’ennemis naturels et de pesticides, le choix des produits phytopharmaceutiques doit prendre en compte les effets éventuels sur la faune auxiliaire. En effet, la lutte intégrée prône l’utilisation de techniques appropriées et de manière aussi compatible que possible. Il convient dès lors de déterminer un traitement pesticide suffisamment efficace pour réduire les populations du ravageur à un niveau permettant de les maintenir sous le seuil économique de dégâts avec l’aide complémentaire des auxiliaires. La sélectivité du traitement est obtenue par le choix de la matière active et par les quantités employées.
De nombreuses recherches entreprises dans le domaine de l’évaluation des effets de pesticides sur les arthropodes utiles ont été effectuées dans le cadre d’un groupe de travail de l’Organisation internationale de lutte biologique (OILB / IOBC) en vue de la sélection de produits phytopharmaceutiques utilisables dans la lutte intégrée. Des méthodes de référence y sont proposées permettant de définir la toxicité des pesticides à l’égard de diverses espèces auxiliaires (Hassan et al. 1994). Les résultats présentés dans cet article s’inscrivent dans ce contexte. Leur objectif consistait à déterminer les effets létaux et sublétaux de quatre produits commerciaux utilisés communément dans la lutte contre les pucerons en cultures maraîchères de plein champ sur l’auxiliaire Episyrphus balteatus De Geer [Diptera : Syrphidae]. Les effets de ces formulations à des doses réduites furent évalués en conditions de laboratoire afin de déterminer la concentration qui autorise une bonne protection de la culture tout en augmentant la sélectivité du traitement.
Matériel et méthodes
Des tests pesticides furent réalisés sur le couple E. balteatus – Megoura viciae Buckton [Homoptera : Aphididae] afin de déterminer les effets létaux et chroniques de deux formulations aphicides : Pirimor® {WG, 50 % m.a. : pirimicarbe [2-dimethylamino-5,6-dimethylpyrimidin-4-yl dimethylcarbamate]} et Karate® {WG, 2,5 % m.a. : lambda-cyhalo-thrine [alpha-cyano-3-phenoxybenzyl 3-(2-chloro-3,3,3-trifluoroprop-1-enyl)-2,2-dimethylcyclopropane carboxylate]} et de deux fongicides : Ronilan® {SC, 500 g m.a. : vinclozoline [3-(3,5-dichlorophenyl)-5-methyl-5-vinyloxazolidine-2,4-dione]} et Quintalic® {SC, m.a. : iprodione 175 g L-1 [3-(3,5-dichlorophenyl-N-isopropyl-2,4-dioxoimidazoline-l-carboximide]; carbendazime 87,5 g L-1 [methyl benzimidazol-2-ylcarbamat]}. La méthode utilisée consistait à mettre en contact les insectes et les pesticides en trempant les feuilles de fève des marais (Vicia faba L. subsp. major) dans une bouillie aqueuse à la concentration correspondant à la dose recommandée (Hassan 1985). Trois concentrations furent sélectionnées pour chacune des matières actives testées : la concentration de la bouillie correspondant à la dose recommandée (Anonyme 1996) pour la culture de fève des marais (100 %), la moitié (50 %) et le dixième (10 %) de cette concentration. Pour les deux insecticides, une concentration supplémentaire à 2,5 % fut aussi testée. Après un séchage d’une heure, les feuilles de fève furent ensuite placées individuellement dans une boîte de Pétri (10 cm de diam) pourvue d’aérations. Les effets létaux et chroniques des quatre formulations aux différentes doses furent observés à chaque fois sur 60 individus. Les larves de syrphe du deuxième stade larvaire furent introduites individuellement dans les boîtes en présence de pucerons (M. viciae) et d’une feuille de fève. Après 24 h d’exposition au pesticide, les auxiliaires furent transférés dans de nouvelles boîtes de Pétri exemptes de pesticide et contenant de nouveaux pucerons. Pour chaque formulation testée, l’essai comprenait aussi 60 larves témoins placées en présence de feuilles non traitées. La mortalité larvaire fut suivie durant 5 j après l’exposition. Les durées de développement larvaire et de pupaison furent également observées jusqu’à l’émergence des adultes. Ces derniers furent ensuite pesés et sexés. Les syrphes furent enfin placés par couple dans une petite cage (60 cm X 30 cm X 30 cm), de manière à pouvoir étudier la fécondité des adultes et la viabilité des oeufs durant 21 j. Le nombre d’oeufs et leur viabilité furent observés pendant 7 j.
Élevage de masse du puceron M. viciae
La fève des marais fut utilisée comme plante hôte pour élever M. viciae. Les graines étaient semées et mises à germer dans des bacs de 30 cm X 20 cm, contenant un mélange (1:1) de vermiculite et de perlite. Les fèves furent inoculées de pucerons au stade deux feuilles. Les élevages furent conduits dans des conditions contrôlées (22 ± 2 °C, 60 à 80 % d’humidité relative et photopériode de 16 L:8 O).
Élevage de masse d’E. balteatus
Les adultes d’E. balteatus furent élevés dans des cages (75 cm X 60 cm X 90 cm) constituées d’une armature en bois recouverte de toile nylon « insect proof », à une température de 22 ± 2 °C, une humidité relative entre 60 et 80 % et une photopériode de 16 L:8 O. Des fleurs artificielles étaient utilisées pour nourrir les syrphes adultes. Ces « fleurs » consistaient en un mélange d’eau (1:4), de miel (1:4), de sucre (1:4) et de pollen (1:4) étendu sur un papier filtre. Huit à 14 j après l’émergence des adultes issus des larves exposées aux pesticides, la ponte fut induite par l’introduction de plantes infestées par M. viciae dans les cages d’élevage. Les plantes furent alors placées en incubateur (20 ± 1 °C, 60 à 80 % d’humidité relative et photopériode de 16 L:8 O) jusqu’à l’émergence des larves.
Traitement statistique des résultats
Le traitement statistique des données et les analyses de variance furent réalisés en utilisant le logiciel MINITAB version 12.2. Pour chacun des tests, les produits et les doses furent comparés entre eux quant à la mortalité qu’ils déterminaient. Les taux de mortalité furent corrigés par rapport au taux de mortalité du témoin correspondant à l’aide de la formule d’Abbott (Abbott 1925). Une fécondité cumulée et corrigée fut déterminée pour chacun des traitements. De manière à caractériser au mieux l’impact d’un pesticide sur la biologie de l’insecte, la viabilité des oeufs (E) et la durée de développement (D) furent également mesurées. Les taux de mortalité cumulés et corrigés, la durée de développement larvaire, la masse des syrphes, ainsi que le taux de fécondité moyen furent comparés en effectuant des analyses de la variance (niveau de signification a = 5 %) éventuellement suivies par des comparaisons de moyennes en utilisant le test de Tukey (T). Enfin, le sex ratio, après transformation angulaire, fut analysé à l’aide d’un test d’égalité de proportions (Dagnelie 1975).
Résultats
Impact sur la mortalité des larves
Karate®
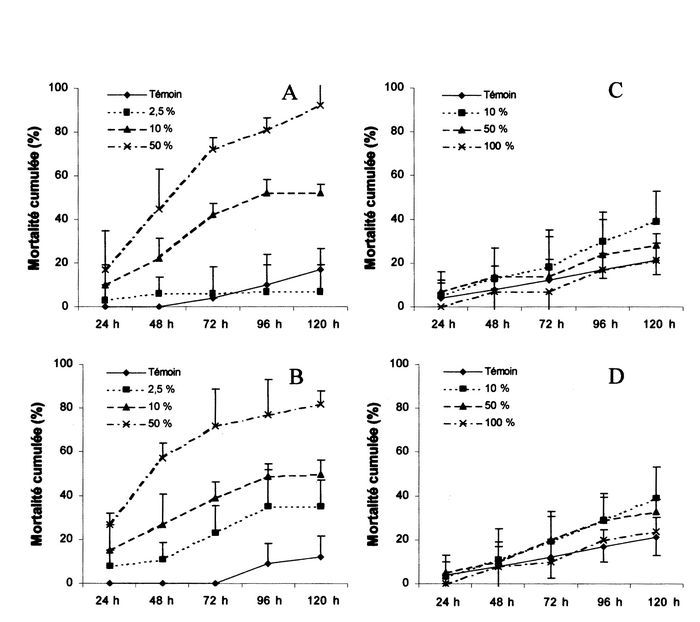
L’analyse de la variance montre que la mortalité cumulée varie significativement en fonction des doses (F = 79,16 ; d.l. = 3, 32 ; P < 0,001) et du temps (F = 6,22 ; d.l. = 4, 32 ; P = 0,001). La concentration 100 % en Karate® a provoqué la mort de l’ensemble des auxiliaires après 24 h. À l’issue de cette même durée, le traitement à la concentration 2,5 % de Karate® a engendré une mortalité significativement inférieure aux traitements 100 et 50 % (respectivement : T = 16,64 ; P < 0,001 et T = 5,27 ; P = 0,021) (Fig. 1 A). De plus, la mortalité induite par Karate® 10 et 50 % fut significativement inférieure (respectivement : T = 14,15 ; P < 0,001 et T = 11,37 ; P = 0,001) à celle provoquée par le traitement Karate® 100 %. Par la suite, et, jusqu’à la fin de l’expérimentation, la mortalité cumulée causée par le traitement Karate® 2,5 % demeura significativement inférieure (T > 6,57 ; P < 0,01) à la mortalité cumulée induite par Karate® 100 %. Après 48 h, une mortalité cumulée significativement inférieure (T = 7,12 ; P = 0,002) au traitement Karate® 100 % fut également observée pour la dose 10 %. En dépit du taux de mortalité évoluant de façon très différente selon le traitement (Fig. 1 A), aucune autre différence significative (T < 3,96 ; P < 0,055) ne fut relevée au cours de l’expérimentation. À l’issue des 5 j, les traitements 2,5, 10 et 50 % auront provoqué des mortalités corrigées cumulées respectives de 10, 68 et 95 %.
Pirimor®
Les concentrations de 100 % en Pirimor® et en Karate® ont provoqué la mort de l’ensemble des larves d’E. balteatus en 24 h. L’analyse de la variance révèle que la mortalité cumulée causée par les traitements au Pirimor® varie de manière significative en fonction des facteurs « temps » (F = 3,79 ; d.l. = 4, 32 ; P = 0,012) et « doses » (F = 42,73 ; d.l. = 3, 33 ; P < 0,001). Après 24 h et 48 h, les traitements Pirimor® 2,5 et 10 % ont engendré une mortalité cumulée significativement inférieure (respectivement T > 5,97 ; P < 0,013 et T > 4,98 ; P < 0,027) à celle engendrée par la dose 100 %. Ici également, bien que le taux de mortalité évolue de façon très différente selon le traitement (Fig. 1 B), aucune autre différence significative (T < 3,96 ; P > 0,054) ne fut notée au cours de l’expérimentation. À la fin de l’essai, les traitements 2,5, 10 et 50 % auront provoqué des mortalités corrigées cumulées de 33, 67 et 82 % respectivement.
Figure 1
Mortalité cumulée des larves d’E. balteatus en fonction des différentes concentrations de Karate® (A), de Pirimor® (B), de Ronilan® (C) et de Quintalic® (D).
Ronilan®
La mortalité cumulée engendrée par les différents traitements à base de Ronilan® varie significativement (F = 6,32 ; d.l. = 4, 23 ; P = 0,001) en fonction du temps, mais aucun effet significatif (F = 0,72 ; d.l. = 2, 25 ; P = 0,497) n’a pu être mis en évidence en fonction des doses. Les taux de mortalité cumulés de l’ensemble des traitements à l’issue de l’expérience sont en outre très semblables et peu sensiblement supérieurs à celui du groupe témoin (Fig. 1 C).
Quintalic®
L’analyse de la variance appliquée aux taux de mortalité corrigés obtenus pour les traitements Quintalic® révèle l’existence d’un effet des doses (F = 15,36 ; d.l. = 2, 23 ; P < 0,001) et d’un effet du temps (F = 17,45 ; d.l. = 4, 25 ; P = 0,001). L’effet des doses ne se manifeste qu’après 72 h où le traitement 50 % a engendré une mortalité corrigée cumulée significativement supérieure (T = 4,19 ; P = 0,049) au traitement 100 %. Par la suite, une mortalité cumulée significativement supérieure est encore observée après 120 h pour les traitements 10 et 50 %, par rapport au traitement 100 % (respectivement : T = 9,92 ; P = 0,005 et T = 6,73 ; P = 0,014). Après 5 j, les traitements Quintalic® 10, 50 et 100 % auront induit des mortalités corrigées cumulées de respectivement 15, 15 et 4 % (Fig. 1 D).
Impact sur la durée de développement larvaire
Pirimor® et Karate® appliqués à 100 % de la dose recommandée ont engendré une mortalité totale chez les larves (Tableau 1). L’analyse de la variance appliquée à la durée de développement larvaire des individus ayant subi les différents tests démontre la présence d’un effet des doses pour les formulations Karate®, Pirimor® et Ronilan® (respectivement : F = 10,46 ; d.l. = 3, 43 ; P < 0,001 ; F = 5,60 ; d.l. = 3,.81 ; P = 0,002 et F = 12,42 ; d.l. = 3, 120 ; P < 0,001). Le traitement Karate® 10 % a, en moyenne, allongé la durée de développement de 4 j par rapport au témoin et aux autres doses. Cet allongement est hautement significatif par rapport à Karate® 2,5 % et au témoin (respectivement : T = 4,89 ; P < 0,001 et T = 5,31 ; P < 0,001). Les larves exposées au traitement Pirimor® 10 % ont mis en moyenne 17,8 % de temps en plus que le groupe témoin pour accomplir leur développement; cet allongement est statistiquement significatif (T = 3,53 ; P = 0,004). Aux doses de 50 et 2,5 %, Pirimor® n’a eu aucun impact significatif sur la durée de développement (T < 0,99 ; P = 0,755). Ronilan® concentré à 100 % a, en moyenne, raccourci significativement (T = 4,11 ; P < 0,001) la durée de développement larvaire de près de 2 j par rapport au témoin et au traitement Ronilan® 50 %, et d’un jour et demi par rapport à la dose 10 %. Enfin, la formulation Quintalic® n’a produit aucun effet significatif sur la durée de développement larvaire (F = 0,88 ; d.l. = 3, 95 ; P = 0,512).
Impact sur la masse des adultes
Aucun traitement n’engendra d’effet significatif sur la masse des femelles d’E. balteatus (F < 2,58; d.l. = 3, 14; 3, 41; 3, 47 et 3, 53 ; P > 0,067). Par contre, les mâles présentèrent une masse moyenne significativement différente selon la dose appliquée pour les formulations Ronilan® et Pirimor® (respectivement : F = 4,05 ; d.l. = 2, 41 ; P = 0,029 et F = 4,54 ; d.l. = 3, 26 ; P = 0,008). Ainsi, la masse moyenne des mâles exposés au Pirimor® 10% fut statistiquement supérieure (T = 2,79 ; P = 0,025) à celle des mâles exposés à la dose 2,5 % (Tableau 1). Cependant, aucune différence significative ne fut observée entre ces deux traitements et le groupe témoin (T < 1,98 ; P > 0,136). Ronilan® 100 % réduisit significativement (T = 3,32 ; P = 0,001) la masse moyenne des mâles par rapport au traitement Ronilan® 10 %. Par ailleurs, aucune différence significative (T < 2,57 ; P > 0,065) ne fut observée entre les trois doses testées et le témoin.
Impact des pesticides sur le sex ratio
À l’exception des syrphes exposés à 100 % de la dose recommandée de Ronilan® (uobs = 2,18 et P = 0,028) aucun des sex ratios ne fut significativement différent (uobs< 1,56 et P > 0,119) de celui du témoin relatif, ni des autres traitements.
Tableau 1
Effet de différentes doses de traitements insecticides et fongicides sur les paramètres biologiques du syrphe E. balteatus
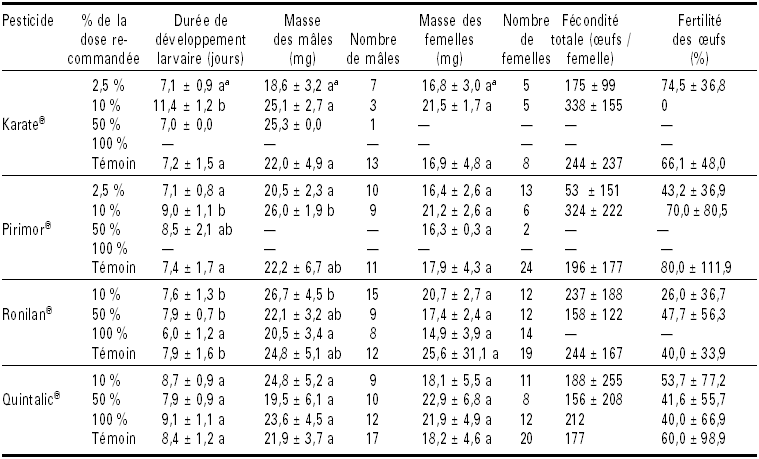
a Dans une même colonne et pour un même produit, les moyennes suivies d’une même lettre ne sont pas significativement différentes (comparaisons selon la méthode de Tukey, au seuil de signification de 5 %).
Discussion
Les insecticides Pirimor® et Karate® sont considérés, à la dose recommandée et en conditions de laboratoire, comme toxiques vis-à-vis d’E. balteatus. La première formulation, très toxique, tue dans les 72 premières h alors que la deuxième agit plus lentement sur la mortalité, sur la fécondité et sur la viabilité des larves (Jansen 1998). D’autres essais, réalisés cette fois en champ, confirment ces résultats. Ainsi Niehoff et Poehling (1995) ont remarqué que le pirimicarbe, à la dose recommandée (100 g m.a. ha-1) et même à 50 % de cette dose, est efficace contre les pucerons mais aussi, malheureusement, contre les syrphes. Par contre, il semble y avoir une certaine sélectivité avec une dose de 25 g m.a. ha-1. Le même auteur estime alors qu’il existe une possibilité d’augmenter la sélectivité en réduisant les dosages, d’autant plus que le taux de mortalité des prédateurs diminue plus vite que celui des ravageurs lorsque les concentrations en pesticide sont réduites. De plus, des résultats obtenus par Gendrier et Reboulet (1992) ont mis en évidence une toxicité modérée à forte du Pirimor® pour les populations de syrphes. Cependant, ces auteurs observent une reconstitution de ces populations en moins de 20 j. Karate® est considéré comme très toxique pour les syrphes, une application entraîne une forte diminution des effectifs (Gendrier et Reboulet 1992). Mises à part ces données, il existe peu d’informations détaillées concernant la toxicité de ces aphicides sur les larves des auxiliaires. En outre, à notre connaissance, aucun effet éventuel des formulations fongicides Ronilan® et Quintalic® n’a été observé. Néanmoins, Branquart et al. (1992) ont observé une mortalité importante chez certains autres groupes d’auxiliaires, dont les coccinelles, suite à un traitement réalisé avec Ronilan®.
Pour les traitements aphicides, l’essentiel de la mortalité est observée les 3 premiers j suivant l’exposition et lors de la nymphose. Les doses de 10 et 50 % de Karate® et de Pirimor® engendrent toutes deux une mortalité importante des syrphes (de 50 à 100 % à l’issue de l’expérience). Les survivants sont plus nombreux pour le traitement au Karate®. Un certain degré de sélectivité vis-à-vis des larves d’E. balteatus apparaît à la concentration 2,5 % Pirimor® alors qu’à cette même concentration, l’effet toxique de Karate® devient insensible.
Concernant les deux fongicides testés, nos résultats montrent que seul Quintalic® possède un effet sur la mortalité des larves d’E. balteatus. De façon inexpliquée, ce sont les bouillies les moins concentrées qui auront causé les mortalités les plus importantes, mais sans signification par rapport au témoin. De façon inexpliquée également, la durée de développement d’E. balteatus est fortement allongée suite à l’exposition à Karate® 10 % alors que les traitements 2,5 et 50 % n’auront produit aucun effet décelable. Les quatre pesticides n’ont eu aucun impact sur la masse des femelles d’E. balteatus. Toutefois, en ce qui concerne les syrphes mâles, il existe un lien direct entre la réduction de leur masse et la concentration en Ronilan®.
L’exposition aux pesticides agit également sur les paramètres de reproduction des syrphes : fécondité totale et taux d’émergence s’en trouvent réduits. Ces observations recoupent celles de Jansen (1998) et de Niehoff et Poehling (1995). Un taux d’émergence nul est observé même après avoir dilué 10 fois une bouillie contenant du Karate® à la dose recommandée. Ces résultats confirment les observations de Jansen (1998) : la fécondité des syrphes exposés à Karate® 50 % est inférieure au témoin et la viabilité des oeufs nulle.
Il ressort de cette étude que les deux aphicides testés menacent potentiellement l’efficacité du contrôle biologique par leur toxicité directe et chronique qui détruit les larves de l’auxiliaire E. balteatus et menace les générations futures en réduisant la fécondité des femelles. Jansen (1998) avait déjà attiré l’attention sur cette dramatique réduction de l’efficacité de prédation d’E. balteatus lors de traitements avec Karate® et Pirimor® en conditions de plein champ. Quintalic® appliqué à la dose recommandée possède, en outre, un effet néfaste sur la survie des populations de syrphes. Des essais ultérieurs en champ seront nécessaires pour confirmer ces hypothèses.
Parties annexes
Remerciements
Les auteurs tiennent à remercier le Ministère Wallon de l’Agriculture et tout spécialement Monsieur le Ministre Happart pour le financement de cette étude qui fait partie d’un programme de recherche intitulé « Évaluation et utilisation de l’entomofaune utile en cultures maraîchères de plein champ ».
Références
- Abbot, W.S. 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18 : 265-267.
- Anonyme. 1996. Liste des pesticides à usage agricole agréés. Ministère des classes moyennes et de l’Agriculture, Bruxelles. 504 pp.
- Branquart, E., J.L. Hemptinne, B. Adam et C. Gaspar. 1992. Production industrielle d’insectes entomophages. Faculté universitaire des Sciences agronomiques de Gembloux, Gembloux, Belgique. pp. 1-6.
- Dagnelie, P. 1975. Théorie et méthodes statistiques. Tome 2 : Les méthodes d’inférence statistique. Les Presses agronomiques de Gembloux, Gembloux, Belgique. 463 pp.
- Gendrier, J.P. et J.N. Reboulet. 1992. Choix de produits phytosanitaires en vergers (1992). Phytoma - La Défense des végétaux 438 : 26-30.
- Hassan, S.A. 1985. Standard methods to test the side-effects of pesticides on natural enemies of insects and mites developed by the IOBC WPRS working group «Pesticides and beneficial organisms». Oepp/Eppo Bull. 15 : 214-255.
- Hassan S.A., F. Bigler, H. Bogenschütz, J. Brun, J. Calis, J. Coremans-Pelseneer, C. Duso, A. Grove, U. Heimbach, N. Heyler, H. Hokkanen, G.B. Lewis, F. Mansour, L. Moreth, L. Polgar, L. Samsoe-Petersen, B. Sauphanor, A. Staübli, G. Sterk, A. Vainio, M. Veire, G. Viggiani et H. Vogt. 1994. Results of the sixth joint pesticide testing programme of the IOBC/WPRS-working group “Pesticides and beneficial organisms”. Entomophaga 39 : 107-119.
- Jansen, J.P. 1998. Side effects of insecticides on larvae of the aphid specific predator Episyrphus balteatus (De Geer) (Dipt. Syrphidae) in the laboratory. Meded. Fac. Landbouwwet. Univ. Gent. 63 : 585-592.
- Niehoff, B. et H.M. Poehling. 1995. Population dynamics of aphids and syrphid larvae in winter wheat treated with different rates of pirimicarb. Agric. Ecosyst. Environ. 52 : 51-55.
Liste des figures
Figure 1
Mortalité cumulée des larves d’E. balteatus en fonction des différentes concentrations de Karate® (A), de Pirimor® (B), de Ronilan® (C) et de Quintalic® (D).
Liste des tableaux
Tableau 1
Effet de différentes doses de traitements insecticides et fongicides sur les paramètres biologiques du syrphe E. balteatus
a Dans une même colonne et pour un même produit, les moyennes suivies d’une même lettre ne sont pas significativement différentes (comparaisons selon la méthode de Tukey, au seuil de signification de 5 %).