
Résumés
Résumé
Les colorants sont largement utilisés dans les imprimeries, les produits alimentaires, cosmétiques et cliniques, mais en particulier dans les industries textiles pour leur stabilité chimique et la facilité de leur synthèse et leur variété de couleurs. Cependant, ces colorants sont à l’origine de la pollution une fois évacués dans l’environnement. La production mondiale des colorants est estimée à plus de 800 000 t•an-1 et les colorants azoïques sont majoritaires et représentent 60-70 %. Compte tenu de la composition très hétérogène de ces derniers, leur dégradation conduit souvent à la conception d’une chaîne de traitement physique-chimique et biologique assurant l’élimination des différents polluants par étapes successives. Dés études ont montré que plusieurs colorants azoïques sont toxiques et mutagènes et le traitement biologique de ces colorants semble présenter un intérêt scientifique majeur. Les traitements physico-chimiques communs (adsorption, coagulation/floculation, précipitation etc.) sont couramment utilisés pour les effluents industriels. Malgré leur rapidité, ces méthodes se sont avérées peu efficaces compte tenu des normes exigées sur ces rejets. Le traitement biologique constitue une alternative fiable; en effet, plusieurs microorganismes sont capables de transformer les colorants azoïques en sous-produits incolores. Les bactéries dégradent les colorants azoïques en deux étapes : un clivage de liaison azo, par l’intermédiaire de l’azoréductase, suivi d’une oxydation des amines aromatiques formées lors de la première étape. L’azoréduction constitue alors une étape clé du traitement des effluents chargés de ces colorants.
Mots-clés :
- Colorants,
- pollution de l'eau,
- industries textiles,
- biodégradation,
- toxicité
Abstract
Dyes are widely used for industrial, printing, food, cosmetic and clinical purposes as well as textile dyeing because of their chemical stability, ease of synthesis, and versatility. Their stability, however, causes pollution once the dyes are released into the environment in effluents. More than 800,000 tons of dyes are annually produced worldwide, of which 60 to 70% are azo dyes. Considering the heterogeneous composition of these latter dyes, their degradation usually requires a chain of physical, chemical and biological treatments assuring the elimination of different pollutants in successive steps. In addition, some azo dyes are toxic and mutagenic and thus the biological treatment of these dyes is now of major scientific interest. Physical-chemical treatments (adsorption, coagulation/flocculation precipitation, etc.) are usually used for industrial effluents. In spite of their rapidity, these methods have turned out to be ineffective in attaining the standards required for these discharges. As a viable alternative, biological processes are receiving increasing interest owing to their cost effectiveness and their ability to produce less sludge. It has been found that some microorganisms can transform azo dyes into colourless products. Bacterial degradation of azo dyes is often initiated by an enzymatic biotransformation step that involves cleavage of azo linkages with the aid of an azoreductase and an electron donor. As the azoreductase in some microorganisms can catalyze the reductive cleavage of azo groups, they have potential advantages in developing bio-treatment methods of wastewater containing azo compounds.
Keywords:
- Synthetic dyes,
- wastewater pollution,
- textile industries,
- biodegradation,
- toxicity
Corps de l’article
1. Itroduction
Nous vivons dans un monde où tout est coloré, nos vêtements, nos aliments nos produits cosmétiques, pharmaceutiques, etc. Ces colorants sont de plus en plus des colorants de synthèse, en raison de leur facilité de synthèse, de leur rapidité de production et de leur grande variété de couleurs si on les compare aux colorants naturels. Aujourd’hui, les colorants de synthèse constituent une véritable industrie et un capital de la chimie moderne. La diversité structurale des colorants de synthèse dérive à la fois de la diversité des groupements chromophoriques qui les composent (groupements azoïque, anthraquinone, triarylméthane et phtalocyanine) et de la diversité de la technologie d’application (coloration réactive, directe, disperse et de cuve). Les vêtements que nous portons sont fabriqués à partir de fibres textiles colorées à l’aide de différentes teintures leur donnant leur couleur définitive. Nous sommes loin d’imaginer que la fabrication de ces mêmes vêtements engendre une pollution de plus en plus importante et dangereuse pour les citoyens. Ces colorants sont pour la plupart synthétiques et représentent aujourd'hui un large groupe de composés chimiques. La production mondiale de ces colorants de synthèse est évaluée à 800 000 t•an-1. Une partie de ces colorants, approximativement 140 000 t•an-1, sont rejetées lors des étapes de fabrication et coloration des tissus (Ben Mansouret al., 2009a). Les industries textiles, et plus particulièrement les phases de teinture et d’ennoblissement, utilisent principalement des produits chimiques, nuisibles pour la santé, comme certains colorants azoïques cancérogènes, et engendrent une pollution des eaux de surface et des nappes phréatiques.
L'hétérogénéité de la composition des produits utilisés pour teindre fait qu’il est extrêmement difficile d’atteindre des niveaux de pollution satisfaisant les seuils imposés par les normes environnementales, après traitement par les techniques utilisées.
Parmi les nombreuses familles de colorants synthétiques, les colorants azoïques sont les plus largement utilisés (60 à 70 %). Ces colorants sont très stables et relativement peu biodégradables. En raison de leurs effets toxiques et/ou carcinogènes, les fabricants et les industries utilisant ces colorants tentent cependant de réduire l’incidence négative de ces molécules sur l’environnement. La stratégie générale consiste, d’une part, à améliorer le pouvoir colorant intrinsèque des molécules et leur fixation chimique sur la matrice receveuse afin de réduire les doses employées, et, d’autre part, à augmenter leur dégradation. L’évaluation des risques liés à ces molécules colorantes repose sur une approche globale ne permettant pas de situer avec exactitude la nature des toxicités induites. Si les molécules colorantes ne sont pas elles-mêmes nécessairement toxiques à l’origine, leurs dérivés issus des processus de biodégradation pourraient l’être. Ce constat nous a amenés à orienter cette revue (i) sur les colorants : leur définition, leur classification, leurs propriétés chimiques et leur(s) toxicité(s); (ii) sur les méthodes de traitement des rejets de colorants et notamment sur les méthodes biologiques de traitement.
2. Les colorants
2.1 Introduction
L’homme a mis des couleurs dans sa vie dès les débuts de son aventure : peintures rupestres d’Altamira et de Lascaux, céramiques mésopotamiennes, vêtements des tombes égyptiennes, décors corporels des populations primitives, etc. Il a d’abord utilisé les pigments des « terres colorées », puis ceux des fibres végétales et animales.
L’industrie des colorants constitue aujourd’hui un domaine capital de la chimie. Les colorants sont employés pour l’impression et la teinture des fibres textiles, des papiers, des cuirs, des fourrures, des bois, des matières plastiques et des élastomères. Ils servent aussi à préparer des peintures, des encres d’imprimerie, des vernis et, comme additifs, à colorer des produits alimentaires et pharmaceutiques. Ils sont utilisés dans l’industrie des cosmétiques, la coloration des métaux (aluminium anodisé), la photographie (sensibilisateurs), la biologie (coloration des préparations microscopiques), les indicateurs colorés, et certains d’entre eux sont employés en thérapeutique (antiseptiques, antimalariques, etc.).
On distingue deux grandes familles de colorants : les colorants naturels (extraits de matières minérales ou organiques) et ceux issus de la synthèse chimique.
Les premiers colorants employés par l’Homme semblent avoir été d’origine minérale (terres colorées). Quand celui-ci a maîtrisé la technique du tissage, il s’est servi de teintures d’origine végétale ou animale.
2.2 Les colorants synthétiques
Les colorants de synthèse ont progressivement supplanté les colorants naturels. Les recherches, menées depuis le milieu du XIXe siècle, ont débouché sur la fabrication de très nombreuses familles de colorants, où l’on trouve souvent des imitations de la structure chimique des colorants naturels. Cette recherche a joué également un rôle important dans l’essor de la chimie organique et dans la compréhension de la nature des molécules.
La mauvéine, le premier colorant de synthèse, a été découverte par hasard par William Henry Perkin en 1856. Elle a été obtenue à partir de l’aniline (tirée du goudron de houille) par action de l’acide sulfurique en présence de bicarbonate de potassium et a permis de teindre la soie en violet. Les premiers colorants dits « azoïques » furent découverts en Grande-Bretagne en 1860. Ils évincèrent rapidement les colorants à base d’aniline, dont la résistance à la lumière était faible. Mais c’est à l’industrie allemande (Badische Anilin und Soda Fabrick : BASF) que revient la contribution la plus importante à l’essor de l’industrie des colorants.
2.3 Les colorants textiles : définitions et structures
Un colorant doit posséder, outre sa couleur propre, la propriété de teindre. Cette propriété résultant d’une affinité particulière entre le colorant et la fibre est à l'origine des principales difficultés rencontrées lors des traitements. En effet, selon le type d’application et d’utilisation, les colorants synthétiques doivent répondre à un certain nombre de critères afin de prolonger la durée de vie des produits textiles sur lesquels ils sont appliqués : résistance à l’abrasion, stabilité photolytique des couleurs, résistance à l’oxydation chimique (notamment les détergents) et aux attaques microbiennes.
L'affinité du colorant pour la fibre est particulièrement développée pour les colorants qui possèdent un caractère acide ou basique accentué. Ces caractéristiques propres aux colorants organiques accroissent leur persistance dans l’environnement et les rendent peu disposés à la biodégradation (Pagga et Brown, 1986).
Les matières colorantes se caractérisent par leur capacité à absorber les rayonnements lumineux dans le spectre visible (de 380 à 750 nm). La transformation de la lumière blanche en lumière colorée par réflexion sur un corps, ou par transmission ou diffusion, résulte de l'absorption sélective d'énergie par certains groupes d'atomes appelés chromophores, la molécule colorante étant le chromogène (Figure 1). Plus la facilité du groupe chromophore à donner un électron est grande, plus la couleur sera intense (groupes chromophores classés par intensité décroissante dans le tableau 1). D'autres groupes d'atomes du chromogène peuvent intensifier ou changer la couleur dû au chromophore : ce sont les groupes auxochromes (Figure 1).
Figure 1
Exemples des groupes chromophores et auxochromes des colorants de types azoïques et anthraquinones.
Example of the chromophoric and auxochromic groups of azo dyes and anthraquinone.
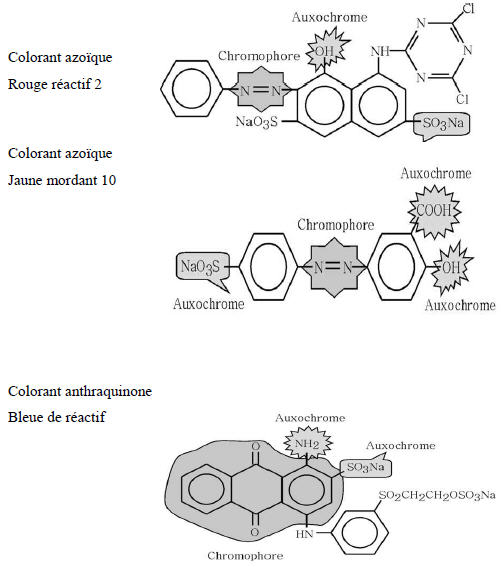
Les chromophores sont des groupes aromatiques (électrons π délocalisés), conjugués (liaisons π), comportant des doublets non liants (électrons n) ou des complexes de métaux de transition. Les colorants diffèrent les uns des autres par des combinaisons d'orbitales moléculaires. La coloration correspond aux transitions possibles après absorption du rayonnement lumineux entre ces niveaux d'énergie propres à chaque molécule (Capon et al., 1999).
De manière générale, les colorants consistent en un assemblage de groupes chromophores, auxochromes et de structures aromatiques conjuguées (cycles benzéniques, anthracène, pérylène, etc.). Lorsque le nombre de noyaux aromatiques augmente, la conjugaison des doubles liaisons s'accroît et le système conjugué s'élargit. L'énergie des liaisons π diminue tandis que l'activité des électrons π ou n augmente et produit un déplacement vers les grandes longueurs d'onde. De même, lorsqu'un groupe auxochrome donneur d'électrons (amino, hydroxy, alkoxy, etc.) est placé sur un système aromatique conjugué, ce groupe se joint à la conjugaison du système π, la molécule absorbe dans les grandes longueurs d'onde et donne des couleurs plus foncées (Christie, 2001; Seyewetz et Sisley, 1896; Welham, 2000; Zhenwanget al., 2000).
2.4 Classification des colorants
Contrairement à l’usage établi en chimie organique, la terminologie employée dans le domaine des colorants n’obéit à aucune règle absolue. Une classification rationnelle des matières colorantes organiques présente de grandes difficultés. Certains auteurs regroupent ces colorants d’après leur constitution chimique, en tenant compte de la nature des chromophores qui les composent : groupement azoïque, anthraquinone, triarylméthane et phtalocyanine (Seyewetz et Sisley, 1896); d’autres les regroupent d’après les diversités des technologies d’application : colorants anioniques (acides, à mordant-acide, directs, réactifs, de cuve, colorants au soufre) ou cationiques (basiques) (Venkataraman, 1901).
Les noms commerciaux rappellent les nuances, leur emploi principal ou le nom de leur inventeur. Les chiffres et les lettres qui suivent parfois le nom précisent la nuance ou certaines propriétés comme la résistance à la lumière ou à divers agents.
2.4.1 Classification chimique
La classification des colorants selon leur structure chimique repose sur la nature du groupe chromophore (Tableau 1). Les colorants azoïques faisant l'objet de cette étude seront traités plus en détail dans la section 2.5.
Tableau 1
Principaux groupes chromophores et auxochromes classés par intensité croissante.
Table 1. Principle chromophoric and auxochromic groups classified by increasing intensity.

Les colorants anthraquinoniques sont, d’un point de vue commercial, les plus importants après les colorants azoïques. Leur formule générale dérivée de l'anthracène montre que le chromophore est un noyau quinonique sur lequel peuvent s'attacher des groupes hydroxyles ou amino.
Les colorants indigoïdes tirent leur appellation de l’indigo dont ils dérivent. Ainsi, les homologues sélénié, soufré et oxygéné du bleu indigo provoquent d’importants effets hypsochromes avec des coloris pouvant aller de l’orange au turquoise.
Les colorants xanthènes, dont le composé le plus connu est la fluorescéine, sont dotés d'une intense fluorescence. Peu utilisés en tant que teinture, leur usage est bien établi comme marqueurs lors d'accidents maritimes ou comme traceurs d'écoulement pour des rivières souterraines, des flux de rejets, etc.
Les phtalocyanines ont une structure complexe basée sur l'atome central de cuivre. Les colorants de ce groupe sont obtenus par réaction du dicyanobenzène en présence d’un halogénure métallique (Cu, Ni, Co, Pt, etc.).
Les colorants nitrés et nitrosés forment une classe de colorants très limitée en nombre et relativement ancienne. Ils sont actuellement encore utilisés, du fait de leur prix très modéré lié à la simplicité de leur structure moléculaire caractérisée par la présence d’un groupe nitro (-NO2) en position ortho par rapport à un groupement électrodonneur (hydroxyle ou groupes aminés).
2.4.2 Classification tinctoriale
Si la classification chimique présente un intérêt pour le fabricant de matières colorantes, le teinturier préfère le classement par domaines d’application. Ainsi, il est renseigné sur la solubilité du colorant dans le bain de teinture, son affinité pour les diverses fibres et sur la nature de la fixation. Celle-ci est de force variable selon que la liaison colorant - substrat est du types ionique, hydrogène, de Van der Waals ou covalente. On distingue différentes catégories tinctoriales définies cette fois par les auxochromes.
Les colorants acides ou anioniques : Très solubles dans l’eau grâce à leurs groupes sulfonate ou carboxylate, ils sont ainsi dénommés parce qu’ils permettent de teindre les fibres animales (laine et soie) et quelques fibres acryliques modifiées (nylon, polyamide) en bain légèrement acide. L'affinité colorant - fibre est le résultat de liaisons ioniques entre la partie acide sulfonique du colorant et les groupes amino des fibres textiles.
Les colorants basiques ou cationiques : Classe des colorants porteurs d’ions positifs et reconnus pour leurs nuances brillantes. Les colorants basiques se composent de grosses molécules et ce sont des sels solubles dans l’eau. Ils ont une affinité directe pour la laine et la soie et peuvent être utilisés sur le coton. La solidité des colorants basiques sur ces fibres est très faible. Ces colorants ont bénéficié d’un regain d’intérêt avec l’apparition des fibres acryliques, sur les quelles ils permettent des nuances très vives et résistantes.
Les colorants de cuve sont des colorants insolubles dans l’eau, appliqués sur la fibre après transformation par réduction alcaline en leucodérivés. La teinture se termine par la réoxydation in situ du colorant sous sa forme insoluble initiale. Réputés pour leur bonne résistance aux agents de dégradation (lavage, rayons solaires), les colorants de cuve sont largement utilisés sur le coton, le lin, la rayonne et autres fibres cellulosiques, à l’image de l’indigo pour la teinture des articles jean ou denim.
Les colorants directs contiennent ou sont capables de former des charges positives ou négatives électrostatiquement attirées par les charges des fibres. Ils se distinguent par leur affinité pour les fibres cellulosiques sans application de mordant, liée à la structure plane de leur molécule.
Les colorants à mordants contiennent généralement un ligand fonctionnel capable de réagir fortement avec un sel d'aluminium, de chrome, de cobalt, de cuivre, de nickel ou de fer pour donner différents complexes colorés avec le textile. On peut distinguer deux types :
Colorants à complexe métallifère type 1:1 : colorants ayant un ou des éléments métalliques dans leur structure moléculaire. Requièrent l’utilisation de l’acide sulfurique.
Colorants à complexe métallifère type 1:2 : deuxième génération des colorants acides traités avec des métaux de mordançage tels que le chrome. Ce type de colorant teint les fibres beaucoup plus solidement que les colorants acides courants. Ils sont appliqués en milieu légèrement acide, soit en pH 4,5 à 5.
Les colorants réactifs contiennent des groupes chromophores issus essentiellement des familles azoïque, anthraquinonique et phtalocyanine. Leur appellation est liée à la présence d’une fonction chimique réactive, de type triazinique ou vinylsulfone, assurant la formation d’une liaison covalente forte avec les fibres. Solubles dans l’eau, ils entrent de plus en plus fréquemment dans la teinture du coton et éventuellement dans celle de la laine et des polyamides.
Les colorants développés ou azoïques insolubles, appelés aussi colorants au naphtol, sont formés directement sur la fibre. Au cours d’une première étape, le support textile est imprégné d’une solution de naphtol ou copulant. Les précurseurs de la molécule suffisamment petits pour diffuser dans les pores et les fibres sont ensuite traités avec une solution de sel de diazonium qui, par réaction de copulation, entraîne le développement immédiat du colorant azoïque. Puisque le composé phénolique est dissous dans une solution basique, ces colorants ne sont utilisés que sur les fibres cellulosiques bien que d’autres fibres soient susceptibles d’être teintes en modifiant le procédé.
Les colorants dispersés appelés aussi plastosolubles sont très peu solubles dans l'eau et sont appliqués sous forme d'une fine poudre dispersée dans le bain de teinture. Ils sont en mesure, lors d’une teinture à haute température, de diffuser dans les fibres synthétiques puis de s'y fixer. Les colorants dispersés sont largement utilisés dans la teinture de la plupart des fibres manufacturées, surtout le polyester.
Les colorants au soufre sont insolubles dans l’eau mais appliqués sous forme d’un dérivé soluble après réduction par le sulfure de sodium. Ils sont ensuite réoxydés à leur état insoluble dans la fibre. Les colorants au soufre sont généralement employés sur le coton pour produire des teintes foncées économiques, dont la solidité au lavage et à la lumière va de moyenne à bonne.
2.5 Les colorants azoïques
C’est en 1863 que Mitscherlich a découvert l'azobenzène C6H5-N = N-C6H5, mais c'est Peter Griess qui a effectué les premiers travaux systématiques à partir de 1858 en décrivant la méthode de préparation très générale de ces produits. Les colorants azoïques constituent la famille la plus importante tant sur le plan des applications qui représentent plus de 50 % de la production mondiale de matières colorantes, soit 800 000 tonnes (Bauer et al., 2001; Ganechet al., 1994; O’Neill et al., 1999; Pandeyet al., 2007), que sur celui de la multiplicité des structures étudiées, soit 60 à 70 % des colorants synthétiques (Zollinger, 1987).
Le nombre de colorants azoïques a connu une évolution importante et a atteint, dans les années 90, plus de 10 000 molécules commercialisées. Ces colorants sont impliqués dans un large éventail de domaines : textile, imprimerie, alimentaire, cosmétique et pharmaceutique (Zollinger, 1987). L’industrie textile représente la partie majeure du marché de ces colorants (Galindo, 1998).
On nomme « azoïques » les composés caractérisés par le groupe fonctionnel azo (-N = N-) unissant deux groupements alkyles ou aryles identiques ou non (azoïque symétrique et dissymétrique). Ces structures, qui reposent généralement sur le squelette de l’azobenzène, sont des systèmes aromatiques ou pseudo-aromatiques liés par un groupe chromophore azo (-N = N-).
L'introduction de groupes azo entre deux noyaux aromatiques déplace le spectre d'absorption du benzène vers les grandes longueurs d'onde de telle sorte que la couleur apparaît (effet bathochrome). Le plus simple des colorants azoïques, l'azobenzène, est jaune-orangé. L'introduction de groupes amines ou phénols a également un effet bathochrome, de même que la multiplication des groupes azoïques, aussi peut-on obtenir presque toutes les nuances du spectre. La présence dans un tel édifice de substituants sulfonés, nitrés ou halogénés, accepteurs ou donneurs d’électrons n ou π délocalisables sur le(s) cycle(s) aromatique(s), permet d'augmenter le phénomène de résonance. C'est ainsi que l'on peut jouer sur la couleur et sur les qualités de teinture. En général, plus le système π de la molécule est conjugué, plus la longueur d'onde qu'il absorbera sera grande. Cependant la complexité des molécules diminue la vivacité des nuances.
Les colorants azoïques forment une gamme étendue de nuances (du jaune au bleu, au vert et même au noir) et se rencontrent dans diverses classes tinctoriales : les colorants basiques, acides, directs et réactifs solubles dans l'eau, les azoïques dispersés et à mordant non ioniques insolubles dans l'eau.
À l’issue du procédé de coloration, une quantité non négligeable (10 à 15 %) des colorants engagés se retrouvent dans les eaux usées (Baueret al., 2001). Or ces composés organiques cancérigènes sont réfractaires aux procédés de traitement habituellement mis en oeuvre et sont très résistants à la biodégradation (Pagga et Brown, 1986).
2.6 Le traitement des effluents textiles (chargés en colorants en particulier azoïques)
2.6.1 Les dangers évidents
Eutrophisation : Différents ions peuvent être évacués dans le milieu naturel tels que le phosphate, utilisé comme détergent lors du processus d’ennoblissement (Yusuff et Sonibare, 2004) ou le nitrate libéré sous l’action des microorganismes sur les colorants (Kaushiket al., 2010). Ces ions minéraux introduits en quantité trop importante peuvent devenir toxiques pour la vie piscicole et altérer la production d’eau potable. Leur consommation par les plantes aquatiques accélère la prolifération anarchique de celles-ci et conduit à l’appauvrissement en oxygène par inhibition de la photosynthèse dans les strates les plus profondes des cours d'eau et des eaux stagnantes.
Sous-oxygénation : Lorsque des charges importantes de matière organique sont apportées au milieu via des rejets ponctuels, les processus naturels de régulation ne peuvent plus compenser la consommation bactérienne d'oxygène. Manahan (1994) estime que la dégradation de 7 à 8 mg de matière organique par des micro-organismes suffit pour consommer l'oxygène contenu dans un litre d'eau.
Couleur, turbidité, odeur : L’accumulation des matières organiques dans les cours d'eau induit l’apparition de mauvais goûts, de prolifération bactérienne, d’odeurs pestilentielles et de colorations anormales. Willmottet al. (1998) ont évalué qu’une coloration pouvait être perçue par l’oeil humain à partir de 5 x 10-6 g•L-1. En dehors de l'aspect inesthétique, les agents colorants ont la capacité d'interférer avec la transmission de la lumière dans l’eau, bloquant ainsi la photosynthèse des plantes aquatiques.
2.6.2 Les dangers à long terme
-
La persistance : les colorants organiques synthétiques, en particulier azoïques, sont des composés très résistants à la dégradation biologique naturelle (Pagga et Brown, 1986). Cette persistance est en étroite relation avec leur réactivité chimique :
Les composés insaturés sont moins persistants que les saturés;
Les alcanes sont moins persistants que les aromatiques;
La persistance des aromatiques augmente avec le nombre de substituants;
Les substituants halogènes augmentent la persistance des colorants tels que les groupements alkyles.
Bio-accumulation : Si un organisme ne dispose pas de mécanismes spécifiques, soit pour empêcher la résorption d’une substance telle qu’un colorant, soit pour l’éliminer une fois qu’elle est absorbée, alors cette substance s’accumule. Les espèces qui se trouvent à l'extrémité supérieure de la chaîne alimentaire, y compris l'homme, se retrouvent exposées à des teneurs en substances toxiques pouvant être jusqu’à mille fois plus élevées que les concentrations initiales dans l'eau.
Sous-produits de chloration (SPD) : Le chlore utilisé pour éliminer les microorganismes pathogènes réagit avec la matière organique pour former des trihalométhanes (THM) (Santé canada, 1999) dont les concentrations peuvent atteindre plusieurs centaines de mg•L-1. Les SPD sont responsables du développement de cancers du foie, des poumons, des reins et de la peau chez l'homme (Millset al., 1998; Santé Canada, 1999).
2.6.3 Mutagénicité / Carcinogénicité
Une étude, effectuée sur le recoupement des DL50 avec les classifications chimiques et tinctoriales des colorants, démontre que les colorants synthétiques organiques les plus toxiques sont les colorants diazo et cationiques (Zollinger, 1987). Or, le caractère électro-attracteur des groupes azo génère des déficiences électroniques, ce qui rend les azoïques peu disposés au catabolisme oxydatif dans des conditions environnementales aérobies (DEPA, 2000).
La toxicité des azoïques due à l’exposition aux colorants et à leurs métabolites n'est pas un fait nouveau. Dès 1895, l’augmentation du nombre de cancers de la vessie observés chez des ouvriers de l'industrie textile est reliée à leur exposition prolongée aux colorants azoïques (Rehn, 1895). Depuis, les travaux effectués sur ces colorants azoïques ont démontré que ces composés chimiques présentaient des effets cancérigènes pour l'homme et l'animal (Brown et Devito, 1993; Chen, 2006; Combes et Haveland-Smith, 1982; DEPA, 2000; IARC, 1982; Medvedevet al., 1988; Percyet al., 1989; Tsudaet al., 2000) (Tableau 2).
Tableau 2
Colorants azoïques révélés mutagènes et/ou carcinogènes.
Table 2. Azo dyes revealed mutagen and/or carcinogen.

Si la plupart des colorants ne sont pas toxiques directement, une portion significative de leurs métabolites l'est. Leurs effets mutagènes, tératogènes ou cancérigènes apparaissent après dégradation de la molécule initiale en sous-produits d'oxydation. Les effets cancérigènes des composés azoïques s’expriment alors, dans certains cas, indirectement par leurs dérivés aminés (Brown et De Vito, 1993; Chunget al., 1981; Ganesh, 1992; IARC, 1982; Rafiiet al., 1997). La liaison azo est la partie la plus labile de ces molécules et peut facilement se rompre, chez des organismes mammifères incluant l'homme, sous l'action enzymatique d’une azoréductase P 450 exprimée au niveau du foie (Zollinger, 1987) ou d’une azoréductase exprimée par des microorganismes intestinaux strictement anaérobie (Chunget al., 1981; Rafiiet al., 1997), pour se transformer en composés aminés cancérigènes.
Ainsi l’azoréduction peut donc augmenter ou diminuer la toxicité du colorant d’origine. Nous pouvons distinguer quatre cas, ceux du :
colorant toxique à l’origine qui devient plus toxique après métabolisation;
colorant toxique à l’origine qui perd sa toxicité après dégradation;
colorant non toxique à l’origine qui reste non toxique après métabolisation;
colorant non toxique à l’origine qui devient très toxique après métabolisation.
La toxicité des azoïques et de leurs dérivés est accrue par la présence de substituants sur le noyau aromatique, notamment des groupes méthyles (Sandhu et Chipman, 1990), nitro (-NO2) et halogènes, particulièrement le chlore (Guivarch et al., 2004). D’autres substitutions telles que celles des groupes sulfonates (SO3H) permettent d’augmenter, d’une part, l’hydrosolubilité du colorant et sa fixation sur le tissu et, d’autre part, de diminuer la toxicité (mutagénicité) aussi bien des colorants d’origine que des métabolites issus de l’azoréduction. Junget al. (1992) ont comparé la toxicité des molécules de type naphtol avec celle de leurs analogues dépourvues des groupes sulfonates et ont souligné le rôle de ces groupements dans la diminution de la mutagénicité.
Selon la DEPA (2000), l’estimation des risques de cancer impose de fixer une concentration limite de 3,1 µg•L-1 en colorant azoïque dans l’eau potable.
Il faut noter que la mutagénicité des colorants azoïques a été évaluée, dans la plupart du temps, à l’aide du test d’Ames utilisant des souches procaryotes Salmonella typhimurium génétiquement modifiées.
3. Traitement des eaux usées
Le secteur textile fait partie des six branches d’activités générant la moitié des flux industriels de pollution. Les effluents issus de ce secteur peuvent être très colorés et difficiles à traiter. La coloration de ces eaux usées est de plus en plus perçue comme une nuisance importante. La plus grande part des effluents est représentée par l’ennoblissement qui englobe les prétraitements (désencollage, blanchissement), la teinture ou l’impression et les opérations qui confèrent aux fibres textiles des propriétés particulières (Figure 2). La plupart de ces traitements sont des grands consommateurs d’eau (200 L•kg-1 en moyenne). En France, selon la Fédération de l’Ennoblissement Textile (FET), la branche traite 500 000 tonnes de tissu par an avec une production d’eaux usées de l’ordre de 100 millions de m3.
Présentation d’un procédé d’ennoblissement textile (Hao et al. 2000).
Figure 2. Textile finishing process (Hao et al. 2000).

Au cours des différentes étapes du procédé de teinture, des quantités plus ou moins importantes de colorants sont perdues par manque d’affinité avec les surfaces à teindre ou à colorer (Tableau 3) et se retrouvent dans les rejets.
Tableau 3
Taux de fixation sur la fibre textile pour les différentes classes tinctoriales de colorants azoïques (O’Neill et al., 1999; Azbar et al., 2004).
Table 3. Rate of fixation on the textile fiber for the various tinctoriales classes of azo dyes (O’Neill et al., 1999; Azbar et al., 2004).

En effet, les colorants se fixent sur les fibres textiles par des liaisons de type Van der Waals, hydrogène et interaction hydrophobe. La fixation du colorant dépend de la nature du colorant et de sa composition chimique. Le lien très fort fibre - colorant au sein du complexe est renforcé par une interaction électrostatique additionnelle quand le colorant et la fibre présentent des charges opposées (Welham, 2000).
Par exemple, dans des conditions alcalines (pH = 9 - 12), à une concentration en sel de 40 à 100 g•L-1 et à des températures élevées (30 - 70 °C), les colorants réactifs tels que la vinyl-sulfone (R-SO2-CH = CH2) et le chlorotriazinyl forment des liaisons avec la fibre sous cette forme. Cependant, les colorants réactifs subissent dans l’eau une réaction spontanée d’hydrolyse qui diminue l’affinité de ces colorants vis-à-vis de la fibre. Par conséquent, une quantité élevée de colorant mis en oeuvre se décharge dans l’eau usée (Haoet al., 2000).
L'efficacité de fixation change avec la classe du colorant azoïque utilisée, qui est autour de 98 % pour les colorants basiques alors qu’elle ne dépasse pas 50 % pour les colorants réactifs comme le montre le tableau 3.
Le traitement des eaux polluées par ces types de colorants est donc devenu une priorité dans notre monde moderne. La mise au point de méthodes et l’optimisation des procédés existants, qui doivent être aussi efficaces que peu coûteux, font donc l’objet d’un nombre considérable de travaux.
En effet, le traitement des rejets textiles, compte tenu de la composition très hétérogène de ceux-ci, conduira toujours à la conception d’une chaîne de traitement assurant l’élimination des différents polluants par étapes successives. La première étape consiste à éliminer la pollution insoluble par l'intermédiaire de prétraitements (dégrillage, dessablage, déshuilage, etc.) et/ou de traitements physiques ou physico-chimiques assurant une séparation solide - liquide. Les techniques de dépollution intervenant le plus couramment en deuxième étape dans les industries textiles, d'après Haoet al. (2000) et Dos Santoset al. (2007) se divisent en trois types : physiques, chimiques et biologiques.
Les procédés les plus simples et les plus anciens d’élimination des polluants réfractaires aux traitements biologiques sont des méthodes physiques de transfert de masse : en général, la floculation et l’adsorption sur charbon actif. Mais, d’une part, ces méthodes déplacent simplement la pollution dans les grandes quantités de boues ainsi créées et, d’autres part, elles ne sont pas suffisamment actives pour résoudre les problèmes liés à la coloration. La coagulation et la floculation sont, par ailleurs, inefficaces en ce qui concerne les colorants basiques, et la récupération des colorants de cuve par adsorption sur charbon actif est médiocre.
C’est pourquoi les méthodes physiques sont remplacées par des procédés chimiques de destruction. Ces derniers sont basés sur l’oxydation des colorants par des agents chimiques qui sont généralement des systèmes générateurs de radicaux libres, en particulier du radical hydroxyle. Malgré leur rapidité, les méthodes chimiques se sont avérées peu efficaces compte tenu des normes exigées sur les rejets. Ces méthodes ne sont pas universelles pour tous les colorants, elles sont très coûteuses et chargent les rejets finaux en nombreux sous-produits chimiques de réaction.
Il apparaît donc intéressant de mettre au point des traitements alternatifs, notamment par voie biologique, qui ont l’avantage d’être moins coûteux, moins polluants et plus efficaces car plus spécifiques. Les procédés les plus couramment rencontrés seront abordés succinctement dans les paragraphes suivants. Nous prendrons soin d'évoquer à la fois leurs avantages et leurs inconvénients vis-à-vis du traitement des colorants. Enfin, la dégradation des colorants par des bactéries, qui fait l’objet de notre travail, sera abordée plus spécifiquement parmi ces traitements biologiques.
3.1 Méthodes physiques
3.1.1 Filtration sur membrane
La filtration sur membrane pilotée par pression hydraulique se décline en microfiltration, ultrafiltration, nanofiltration et osmose inverse. L’effluent passe à travers une membrane semi-perméable qui retient en amont les contaminants de taille supérieure au diamètre des pores, pour produire un perméat purifié et un concentré qui reçoit les impuretés organiques (Robinsonet al., 2001). Parmi les quatre types de procédés, la nanofiltration et l’osmose inverse sont les plus adaptés à la rétention partielle de la couleur et des petites molécules organiques (Taylor et Jacobs, 1996) et l’osmose inverse reste la plus répandue (Calabroet al., 1990). La nanofiltration s’applique surtout au traitement des bains de teinture de colorants réactifs en agissant comme un filtre moléculaire tandis que la microfiltration retient les matériaux colloïdaux tels que les colorants dispersés ou de cuve grâce à une « membrane écran » (Van Der Bruggenet al., 2003). L’ultrafiltration ne s’applique qu’à la réduction de DCO et des solides en suspension (Anselme et Jacobs, 1996) et ne se montre réellement efficace qu’en combinaison avec la coagulation/floculation.
Ces procédés, limités dans leurs applications, nécessitent des investissements importants (Van Der Bruggenet al., 2003) et le retraitement du concentré est jusqu’à six fois plus cher que celui de l’effluent originel.
3.1.2 Adsorption (sur charbon actif)
Lors de l’adsorption, le colorant est transféré de la phase liquide vers la phase solide. Le charbon activé est le plus communément utilisé pour réduire la couleur, mais cette technique n’est efficace que sur certaines catégories de colorants (cationiques, à mordant, dispersés, de cuve et réactifs (Haoet al., 2000 ; Raghavacharya, 1997; Robinsonet al., 2001). De plus, ces techniques non destructives requièrent des opérations postérieures de régénération et de post-traitement des déchets solides onéreuses. Dans la plupart des cas, les résidus solides sont répandus en décharges et des dispositions particulières doivent être prises à l’égard des composés organiques qui peuvent lixivier avec le temps.
3.1.3 Méthode physico-chimique de coagulation - floculation
Sous le terme de coagulation - floculation, on entend tous les processus physico-chimiques par lesquels des particules colloïdales ou des solides en suspension fines sont transformés par des floculants chimiques en espèces plus visibles et séparables (les flocs). Les flocs formés sont séparés par décantation et filtration puis évacués. Les coagulants inorganiques tels que l’alun sont les plus satisfaisants pour la décoloration des effluents textiles contenant des colorants de cuve et au soufre, mais sont totalement inefficaces pour les colorants réactifs, azoïques, acides et basiques (Haoet al., 2000; Robinsonet al., 2001; Vendevivereet al., 1998). Par ailleurs, la coagulation - floculation ne peut pas être utilisée pour les colorants fortement solubles dans l’eau. Enfin, d’importantes quantités de boue sont formées avec ce procédé : leur réutilisation reste la seule issue mais demande des investissements supplémentaires pour les régénérer.
3.2 Traitements chimiques
Dans la littérature, les techniques chimiques d’oxydation sont généralement appliquées (i) pour le traitement des composés organiques dangereux présents en faibles concentrations, (ii) en prétraitement avant des procédés biologiques, (iii) pour le traitement d’eaux usées chargées de constituants résistant aux méthodes de biodégradation et enfin (iv) en post-traitement pour réduire la toxicité aquatique. Parmi les méthodes de traitement chimique, les procédés d’oxydation avancée (AOP) restent les plus fréquemment utilisés. Ceux-ci sont fondés sur la formation d’une entité radicalaire extrêmement réactive : le radical hydroxyle (•OH) qui possède un temps de vie très court, un potentiel d’oxydation élevé et une forte réactivité vis-à-vis de nombreux composés organiques. Les principaux procédés de production du radical hydroxyle sont présentés.
3.2.1 Quelques procédés d’oxydation avancée (POA)
3.2.1.1. Réactif de Fenton (H2O2/Fe2+)
Fenton avait décrit à la fin du XIXe siècle que le fer ferreux favorisait fortement l’oxydation de l’acide maléique par le peroxyde d’hydrogène en milieu acide (Fenton, 1894). Des travaux ultérieurs ont montré que la combinaison de H2O2 et de Fe2+, nomméE « réactif de Fenton », était un oxydant efficace pour une grande variété de substrats organiques, notamment les phénols, les pesticides, les aromatiques polycycliques et des colorants, en particulier les azoïques (Benitez et al., 2001; De Heridiaet al., 2001; Kondoet al., 2002; Wang et al., 2005). Quarante ans plus tard, Haber et Weiss (1934) identifiaient le radical hydroxyle comme étant l’espèce oxydante de la réaction présentée ci-dessous et communément appelée réaction de Fenton :
La vitesse de décomposition de H2O2 par le Fe(II) augmente lorsque le pH augmente (≤ 5) car, dans cette gamme de pH, la forme prédominante Fe(OH)2+ est beaucoup plus réactive que l’ion Fe2+ (Kuo, 1992).
Les micropolluants organiques, notamment les colorants synthétiques, sont ensuite oxydés par le radical hydroxyle selon une cascade réactionnelle complexe. Les principaux paramètres déterminant la réaction d’élimination d’un micropolluant sont bien connus : temps de contact, température, concentration en peroxyde d’hydrogène et en sulfate de fer et le pH.
L’étude de dépollution des effluents textiles colorés par le procédé Fenton (système Fe2+/H2O2) a été révélée très efficace et plusieurs études ont montré que le taux de minéralisation des colorants synthétiques augmente avec l’augmentation des doses des réactifs et du temps de réaction. Le rapport des réactifs R = [H2O2]/[Fe2+] et le rapport [Fe2+]/[colorant] jouent aussi un rôle important sur la vitesse de dégradation de colorants de départ et sur le taux de minéralisation (Kuo, 1992).
Le procédé Fenton est considéré aujourd’hui comme le procédé le plus utilisé en Tunisie et dans le monde pour le traitement d’effluents industriels textiles, mais ce procédé est limité par le manque de régénération du catalyseur qui nécessite généralement un apport constant en réactifs et qui contraint de ce fait à approvisionner en continu le milieu en peroxyde d’hydrogène coûteux.
3.2.1.2 L’Ozonation (O3)
L’ozone, oxydant puissant, se décompose rapidement en dioxygène et oxygène atomique et doit être produit impérativement « in situ » grâce à l’émission d’une charge électrique sous haute tension dans un courant d’air sec. L’emploi de l’ozone sur les colorants a montré que les effluents chargés réagissent différemment selon leur composition (Alvareset al., 2001; Neamtuet al., 2004; Solozhenkoet al., 1995; Zhanget al., 2004). Les rejets contenant des colorants dispersés et soufrés sont particulièrement difficiles à décolorer (Solozhenkoet al., 1995), alors que ceux chargés de colorants réactifs, basiques, acides et directs le sont assez rapidement. Bien que le pouvoir oxydant de l’ozone soit élevé, une minéralisation complète par ozonation est très difficile (Szpyrkowiczet al., 2001; Tzitziet al., 1994). Le principal désavantage de l'application d’un système d’ozonation aux bains de teinture, outre un investissement et des coûts opératoires élevés imputables à l’énergie électrique consommée et à la maintenance, réside dans la génération de sous-produits de dégradation récalcitrants et toxiques (Alvares et al., 2001; Ince et Tezcanli, 2001). De plus, l'ozonation est limitée par la très faible solubilité de O3 dans l'eau: environ 0,1 mM à 293 K (Lide, 1999) et son transfert de masse est un facteur limitant.
3.2.1.3 Peroxonation (O3/H2O2)
Les radicaux hydroxyles, principaux intermédiaires oxydants de la peroxonation, sont formés par réaction entre l’ozone et le peroxyde d’hydrogène :
Même si ce procédé est plus efficace que l’ozonation pour de nombreux substrats organiques, notamment pour les colorants, son efficacité reste limitée par la vitesse de la réaction entre O3 et H2O2. Ce système est affecté par les mêmes contraintes que l’ozonation (Hernandezet al., 2002; Slokar et Le Marechal, 1998). En revanche, l'ozonation et la peroxonation présentent l'avantage, par rapport aux procédés avec irradiation UV, de pouvoir fonctionner dans une eau à forte turbidité, car le système ne dépend pas de la transmission des rayonnements dans l'effluent.
3.2.1.4 Photolyse de peroxyde d’hydrogène (UV-H2O2)
Sous irradiation ultraviolette, la molécule de peroxyde d’hydrogène subit une coupure homolytique pour conduire à deux radicaux hydroxyles :
La photolyse de H2O2 est plus avantageuse que l’ozonation et la peroxonation et son application est moins complexe, mais son efficacité est moindre en raison du faible coefficient d’extinction de H2O2 dans l’UV. Dans un système à réacteur fermé, le gain en efficacité pour des eaux de forte absorption UV sera limité malgré une hausse de la concentration en peroxyde. De plus, ce réactif est extrêmement instable lorsqu’il est concentré et sa décomposition en eau et en oxygène est fortement exothermique. Par ailleurs, la production des radicaux est affectée par les conditions de milieu telles que la température, le pH, la concentration en H2O2 et la présence de consommateurs de radicaux (Honget al., 1996; Slokar et Le Marechal, 1998).
Selon Shu et Huang (1995), les colorants acides se dégradent plus facilement selon ce type de procédé mais l’efficacité diminue avec l’augmentation du nombre de liaisons azoïques. Cependant, la dégradation des colorants de type réactif jaunes ou verts nécessite un temps de contact très important alors que pour certains autres, comme ceux de type direct, métal - complexe et disperse, la décoloration est très rapide.
3.2.1.5 Photocatalyse de peroxyde d’hydrogène (TiO2/UV/H2O2)
L’efficacité de la dégradation photochimique est considérablement améliorée par l’ajout de catalyseurs semi-conducteurs homogènes ou hétérogènes. Parmi les photocatalyseurs les plus communément rencontrés, l’oxyde de titane (TiO2) présente une stabilité photochimique et une activité photocatalytique favorable au traitement des colorants (Bessekhouadet al., 2003). L’excitation des électrons à la surface du semi-conducteur par les photons UV, d’énergie supérieure à l’énergie d’activation du semi-conducteur crée des trous déficients en électrons dans les couches de valence, aboutissant à la formation du radical hydroxyle et de l’anion superoxyde.
La décoloration et la dégradation des colorants sont gouvernées par les réactions oxydantes dont l’efficacité dépend de la concentration en oxygène. Le principal inconvénient est le manque de connaissance des produits de dégradation générés, les produits finaux pouvant être plus toxiques que les colorants de départ (Wanget al., 2005).
3.2.4 Résumé comparatif des procédés et analyse critique
Bien que les méthodes physiques, chimiques et physico-chimiques soient très rapides, elles se sont avérées très coûteuses et peu efficaces compte tenu des normes exigées. En effet, plusieurs études ont montré que ces méthodes sont, d’une part, efficaces contre certaines des molécules colorantes et pas sur d’autres et, d’autre part, qu’elles chargent le milieu naturel avec des polluants organiques parfois plus toxiques que les molécules d’origines (DOS Santoset al., 2007; Robinsonet al., 2001). Il est à noter aussi que ces méthodes sont très coûteuses et nécessitent un équipement bien sophistiqué (Haoet al., 2000; Robinsonet al., 2001). Ce constat a permis aux chercheurs et aux industriels de s’orienter vers la recherche d’autres méthodes qui soient plus efficaces et moins coûteuses : ce sont les procédés biologiques.
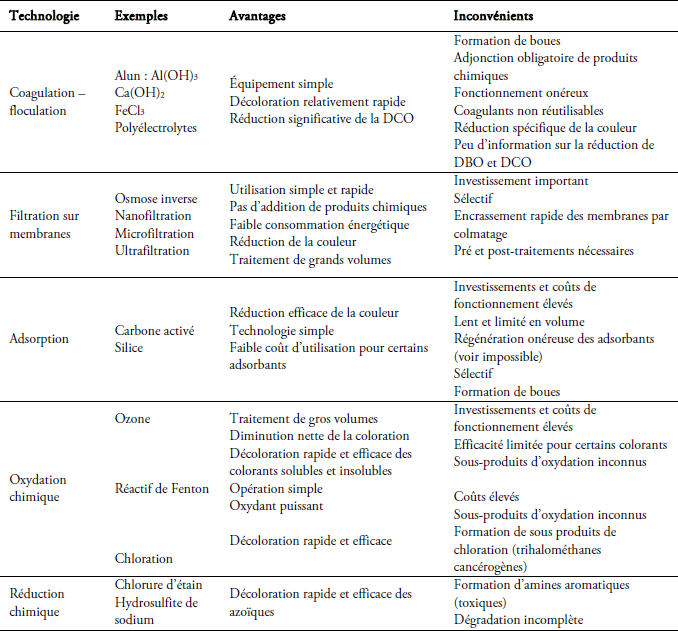
Après ce bref aperçu des principes de fonctionnement des procédés physiques et chimiques de dépollution des rejets de colorants, le tableau 4 résume les types d’application et leurs principaux avantages et inconvénients.
Tableau 4
Comparaison de technologies physiques et chimiques, de dépollution des effluents textiles chargés de colorants synthétiques (Galindo, 1998).
Table 4. Comparative study of physical and chemical technology for the depollution of textile effluents charged by the synthetic dyes (Galindo, 1998).
3.3 Traitements biologiques
3.3.1 Décoloration par les champignons
Les champignons blancs de putréfaction (white-rot fungi) sont capables de dégrader la lignine, structure polymère des plantes. Phanerochaete chrysosporium est le champignon le plus étudié en regard de la dégradation des xénobiotiques tels que les dioxines, les hydrocarbures aromatiques polycycliques (HAP) et autres composés organiques chlorés (Fujianet al., 2001).
La dégradation des colorants par « White-rot fungi » a été rapportée pour la première fois par Glenn et Gold en 1983. Ces auteurs ont étudié l’activité ligninolytique de P. chrysosporium vis-à-vis un certain nombre de colorants polymériques sulfonés. Une étude réalisée par Crippset al. (1990) a montré que P. chrysosporium est capable de dégrader les colorants azoïques sulfonés. Les études menées sur ce type de champignons se sont par la suite multipliées (Tableau 5) sur une gamme plus étendue et plus diversifiée de colorants synthétiques, notamment azoïques (Balan et Monteiro, 2001; Banatet al., 1996; Paszczynskiet al., 1992).
Tableau 5
Quelques études sur la réduction de colorants azoïques par des bactéries aérobie incubées dans des conditions limitées en oxygène.
Table 5. Some studies on the azo dyes reduction by anaerobic bacteria incubated in the limited oxygen conditions.

À côté de P. chrysosporium, d’autres champignons de types « White rot fungi » ont été testés récemment et se sont révélés capables de simplifier les molécules colorantes. Eichlerovaet al. (2005), en testant un ensemble de 30 souches, ont observé l’efficacité de ces microorganismes vis-à-vis des colorants synthétiques, en particulier celle des souches Dichomitus squalents, Ischnoderma resinosum et Pleurotus calyptratus identifiées très actives vis-à-vis de l’orange G et du bleu de rémazole brillant (RBBR).
Harazono et Nakamura (2005) ont montré que la souche Phanerochaete sordida est capable de décolorer un mélange de quatre colorants réactifs : bleu 5, orange 14, rouge 120 et vert 5 avec un pourcentage de décoloration de 90 % après 48 h d’incubation.
L’oxydation des colorants synthétiques (notamment azoïques) par les champignons de putréfaction blanche impliquerait l’expression d’enzymes lignolytiques extracellulaires non spécifiques : les peroxydases telles que la lignine peroxydase (LiP) et la manganèse peroxydase (MnP) ou les phénoloxydases tels que la laccase.
Contrairement aux bactéries, la dégradation des colorants par les champignons est extracellulaire. Le mécanisme d’action des deux enzymes LiP et MnP est similaire : le mécanisme commence par l’oxydation de ces enzymes durant leur cycle catalytique par H2O2, la forme oxydée ainsi formée serait immédiatement réduite à sa forme d’origine par un substrat qui est le colorant azoïque. La différence entre ces deux enzymes est que la LiP oxyde les composés aromatiques phénoliques et non phénoliques, alors que le MnP oxyde le Mn2+ en Mn3+, ce dernier étant responsable de l’oxydation des composés phénoliques uniquement (Banatet al., 1996; Eichlerovaet al., 2005; Fujian et al., 2001; Harazona et Nakamura, 2005).
Les phenoloxydases, divisés en tyrosinases et laccases, sont des oxydoréductases qui peuvent catalyser l'oxydation des composés phénoliques et d'autres composés aromatiques, tels que les colorants (Figure 3), sans utilisation des cofacteurs (Duranet al., 2002).
Figure 3
Mécanisme de dégradation de 4-(4’-sulfophénylazo)-2,6-diméthylephénole par la laccase de Pyricularia oryzae (Chivukula et Renganathan, 1995).
Figure 3. Degradation mechanism of 4-(4’-sulfophenylazo)-2,6-dimethylephenol by laccase from Pyricularia oryzae (Chivukula et Renganathan, 1995).

Paszezynskiet al. (1992) ont étudié et comparé l’efficacité de la dégradation de certains colorants azoïques sulfonés par P. chrysosporium avec des souches d’actinomycètes; ils ont observé que la cinétique de décoloration est beaucoup plus rapide en présence de P. chrysosporium et que les substituants sulfonés limitent la performance des actinomycètes. Ce comportement a été observé également avec des bactéries. En effet, Kullaet al. (1983) ont constaté que les substituants sulfonés (SO3H) inhibent fortement la biodégradation des colorants azoïques.
Bien que les champignons soient les premiers microorganismes identifiés capables de dégrader les colorants synthétiques et qu'ils se soient montrés efficaces dans certains cas comparés aux autres microorganismes, le traitement des rejets textiles chargés en colorants par les champignons pose beaucoup de problèmes. En effet, l’effluent textile n’est pas l’environnement adéquat pour la croissance et la conservation de la biomasse fongique (Robinsonet al., 2001), car le traitement des colorants dans un volume d’eau important (unité de traitement biologique) étant très difficile, il est nécessaire de concentrer les colorants en réduisant la quantité d’eau (Nigam et Marchant, 1995; Nigam et al., 1996).
En outre, l’activité de la lignine peroxydase, meilleure à un pH acide (pH = 4,5 à 5), exige une acidification de l’eau usée qui est généralement très alcaline. Outre un coût d’acidification élevé, cela inhibe la croissance des autres microorganismes utiles, tels que les bactéries, surtout si le traitement est réalisé avec un consortium de microorganismes (Swamy et Ramsay, 1999). Par ailleurs, d'autres polluants de l’eau usée, particulièrement les composés aromatiques, peuvent interférer avec la dégradation fongique des colorants.
3.3.2 Décoloration par les actinomycètes
Les actinomycètes, en particulier l’espèce Streptomyces, produisent les peroxydases extracellulaires qui jouent un rôle primordial dans la biodégradation de la lignine. Ces peroxydases sont impliquées dans l’oxydation de la lignine aboutissant à la production de composés polymériques hydrosolubles. La capacité des actinomycètes à décolorer mais aussi à minéraliser les colorants textiles, notamment azoïques, a été étudiée initialement par trois groupes de chercheurs.
Ballet al. (1989) ont testé 20 souches d’actinomycètes, représentant un large éventail de ce genre, pour leur capacité à décolorer le Poly R. Ces auteurs ont observé que seulement trois souches (Streptomyces badius 252, Streptomyces sp. souche EC22 et Thermomonospora fusca MT800) décolorent significativement le colorant.
Zhou et Zimmermann (1993) ont testé séparément l’aptitude de 159 actinomycètes à dégrader les colorants synthétiques. Cette étude a été réalisée,dans des conditions aérobie sur des effluents textiles similaires contenant séparément des colorants réactifs de structures différentes (le rouge réactif 147 et le bleu réactif 116). Les auteurs ont isolé 83 souches capables de décolorer et de minéraliser ces colorants.
Enfin, un groupe de l’Université de l’Idaho a testé la capacité des microorganismes ligninolytiques, champignons blancs de putréfaction et Streptomycètes, à décolorer et minéraliser des colorants textiles. Dans cette étude, 14 souches de Streptomycètes se sont révélées efficaces sur la dégradation de deux colorants : le Poly B 411 et le Poly R 478. Les auteurs ont suggéré l’implication de peroxydases dans le processus de décoloration (Pastiet al., 1990). Le même groupe (Burke et Crawford; 1998) a partiellement purifié la peroxydase extracellulaire de S. viridosporus T7A dans l’objectif d’identifier la classe de peroxydase responsable de la dégradation des colorants par les Streptomycètes. Les auteurs ont observé que la peroxydase purifiée était similaire et présentait une grande homologie avec la manganèse peroxydase de Phanerochaete chrysosporium.
3.3.3 Décoloration par des algues
L’action décolorante des algues a fait l’objet d’un nombre très limité de travaux. Une étude réalisée par Jinqi et Houtian (1992) a montré que les espèces Chlorella, Oscillatoria et Spirogyra étaient capables de dégrader les colorants azoïques. Leur action décolorante dérive de l’expression d’une azoréductase (enzyme responsable de la fission de la liaison azote-azote) aboutissant à la production des amines aromatiques correspondantes qui sont par la suite complètement oxydées.
3.3.4 Décoloration par les levures
Dans la littérature, les études portant sur la dégradation des colorants azoïques par des levures sont très limitées. Récemment, Ramalhoet al. (2002) ont testé la souche de levure Candida zeylanoides pour réduire des colorants azoïques modèles. En 2004, cette même équipe a pu caractériser l’activité enzymatique responsable de la dégradation des colorants azoïques chez Issatchenkia occidentalis et présenter un an plus tard le système enzymatique d’azoréduction impliqué dans un travail avec Saccharomyces cerevisiae (Ramalho, 2005).
Le nombre de travaux réalisés sur les levures reste très limité en raison de la difficulté à les manipuler et des inconvénients majeurs qu’elles procurent. En effet, outre la difficulté à les cultiver, l’efficacité des levures vis-à-vis des colorants est très faible (la cinétique de décoloration est lente et peut prendre plusieurs dizaines de jours) car les levures nécessitent de s’adapter, c'est-à-dire de s’acclimater avant d’aborder la décoloration proprement dite.
3.3.5 Décoloration par les bactéries
De nombreuses études ont montré la capacité des bactéries à dégrader les colorants. Contrairement aux champignons et aux actinomycètes, qui dégradent les colorants par voie extracellulaire (implication des LiP, MnP, laccases, etc.), les bactéries agiraient plutôt par voie intracellulaire. L’action décolorante dépendrait alors non seulement de l’activité enzymatique cytoplasmique mais aussi de la filtration des molécules à travers la membrane cellulaire.
Dans la littérature, la dégradation complète ou « minéralisation » de colorants azoïques par les bactéries est décrite par la succession de deux étapes essentielles : une azoréduction anaérobie suivie d’une oxydation aérobie des amines aromatiques formées lors de l’étape précédente. L’azoréduction est décrite comme l’étape clé de la minéralisation des colorants, notamment cette étape est suffisante pour la décoloration des molécules.
Certaines souches échappent cependant à cette condition d’anaérobiose et sont capables de réduire les colorants azoïques en présence d’oxygène. On distingue alors deux catégories de bactéries décolorantes selon leur comportement vis-à-vis de l’oxygène : aérobie et anaérobe.
Dans les paragraphes qui suivent nous allons porter une attention plus particulière sur les travaux effectués avec des bactéries du genre Pseudomonas.
3.3.5.1 Dégradation des colorants azoïques par des bactéries dans des conditions limitées en oxygène
La dégradation des colorants azoïques par les bactéries, dans des conditions anaérobie, a été très largement étudiée : on y distingue des bactéries, strictement anaérobies (Bacteroides sp., Eubacterium sp., Clostridium sp., Fusobacterium sp., etc.), anaérobies/aérobies facultatives (Proteus vulgaris, Streptococcus faecalis, etc.) et aérobies (Bacillus sp., Aeromonas hydrophia, Pseudomonas sp., etc.).
Les conditions de dégradation dans la digestion anaérobie sont adaptées à la réduction des colorants azoïques par clivage de la double liaison N = N, appelée azoréduction, entraînant une destruction subséquente des groupes chromophores (celle du système d’électrons π largement délocalisé) mais pas une minéralisation complète. Les amines aromatiques résultantes étant généralement incolores, la réduction azoïque du colorant est aussi désignée dans ce cas par « décoloration ».
Brohm et Frohwein (1937) ont isolé, dès 1937, la première souche bactérienne lactique intestinale capable de réduire les colorants azoïques alimentaires. Comme la formation des amines aromatiques toxiques chez l'homme est un problème majeur, la recherche sur une réduction bactérienne des colorants azoïques a été la plupart du temps concentrée sur l'activité des bactéries anaérobie (facultatives) des intestins des mammifères (Chunget al., 1978; Chunget al., 1981; Chunget al., 1992; Manninget al., 1985; Rafiiet al., 1997; Rafii et Cirniglia, 1993; Walker, 1970).
Horitsuet al. (1977) ont isolé de la boue activée, Bacillus subtilis, la première bactérie non intestinale capable de dégrader les colorants azoïques. Le genre Bacillus a été, par la suite, très largement étudié, à savoir : Bacillus cereus (Wuhrmannet al., 1980), Bacillus subtilis (Yatomeet al., 1991), Bacillus sp. OY1-2 (Suzukiet al., 2001), Bacillus sp. souche SF (Maieret al., 2004). Les études ont aussi concerné d’autres bactéries : Idaka et Ogawa (1978) ont montré en particulier que la souche Aeromonas hydrophila var 24B était capable de dégrader les colorants azoïques. D’autres souches d’Aeromonas hydrophila ont été par la suite testées sur une gamme plus étendue de colorants (Chenet al., 2003; Renet al., 2006).
Les Pseudomonas ont enfin fait l’objet d’un nombre important d’études et ont révélé une grande capacité à simplifier les molécules colorantes (Tableau 5) comme Pseudomonas pseudomallei 13NA (Yatomeet al., 1981), Pseudomonas luteola (Changet al., 2001; Hu, 1994), Pseudomonas putida (Abrahamet al., 2003; Ghorpade et Spencer, 1993); Pseudomonas sp. PR41-1 (Suguiraet al., 1999); Pseudomonas desmolyticum NCIM 2112 (Kalmeet al., 2007); Pseudomonas GM3; Pseudomonas Q3 et Pseudomonas Z1 (Yuet al., 2001). Dix souches de Pseudomonas ont aussi été testées séparément puis en mélange et se sont révélées très actives sur la réduction des colorants azoïques (Fang et al., 2004).
3.3.5.1.1 Azoréduction
On appelle azoréduction le clivage de la double liaison azoïque (-N = N-). Ce phénomène nécessite un transfert de quatre électrons en deux étapes. Dans chaque étape on donne deux électrons au colorant azoïque qui est en fait un accepteur final d’électrons (Dos Santoset al., 2007) :
Cette réaction est catalysée par une enzyme cytoplasmique non spécifique appelée azoréductase. Afin de comprendre si l’action de l’azoréductase était intra- ou extracellulaire, Yu et al. (2001) ont arrêté la réaction de décoloration au bout de 2 h d’incubation en présence de Pseudomonas sp. et ont repris ensuite l’expérimentation séparément, soit avec la biomasse bactérienne, soit avec le milieu de culture. Les auteurs ont observé une faible décoloration (3 %) avec le milieu de culture alors que les colorants disparaissent complètement en présence de biomasse cellulaire. Ils ont conclu, par conséquent, que l’azoréduction est intracellulaire. L’identification des produits issus de l’azoréduction dans le milieu extracellulaire indique que les colorants sont transférés à l’intérieur de la cellule où ils seront réduits par une azoréductase puis les amines dérivées, résistant à l’oxydation, sont rejetées dans le milieu extérieur (Chang et al., 2001; Hsueh et Chen, 2007).
L’azoréductase nécessite, pour son action, la présence d’un cofacteur tel que le NADH, NADPH ou FMN jouant le rôle d’un donneur d’électrons (Figure 4).
Figure 4
Réduction du rouge réactif 22 par Pseudomonas luteola (Chang et al., 2001).
Figure 4. Reduction of reactive red 22 by Pseudomonas luteola (Chang et al., 2001).

Suzuki et al. (2001) sont les premiers à décrire la séquence de gènes codants pour l’azoréductase de Bacillus sp. OY1-2. La protéine de l’azoréductase exprimée par Bacillus sp. OY1-2 est composée de 178 acides aminés constituant alors une nouvelle enzyme de la famille des réductases. Maieret al. (2004) ont identifié l’azoréductase de Bacillus sp. souche SF, enzyme de faible poids moléculaire (62,6 KDa). Les azoréductases isolées de Bacillus subtillis ATCC6633, Bacillus subtillis ISW1214 et Geobacillus stearotherophilus IFO13737 sont formées chacune d’une séquence de 174 acides aminés et présentent des similitudes respectivement de 53,3, 53,9 et 53,3 % avec celle de Bacillus sp. OY1-2 (Suguiraet al., 2006). L’azoréductase isolée de Staphylococcus aureus ATCC 25923, formée de 188 acides aminés, a été comparée à celle de Bacillus sp. OY1-2 et a révélé un degré d’homologie qui ne dépasse pas 32 % (Chenet al., 2005).
On peut constater que, malgré le fait que les azoréductases isolées de différentes souches ne soient pas spécifiques au colorant substrat, ces enzymes ne sont pas tout à fait homologues.
3.3.5.1.2 Influence des paramètres physico-chimiques sur la cinétique de la décoloration
Effet de l’oxygène : De nombreuses études ont montré que l’oxygène est un facteur limitant de la biodégradation des colorants azoïques (Chang et Lin, 2000; Hu, 1994). Selon Changet al. (2001), cet effet est dû à une compétition pour le cofacteur NADH entre la respiration et l’azoréduction. Ces auteurs, en étudiant l’activité azoréductase de l’extrait cellulaire brut issu de P. luteola dans des conditions aérobie, ont prouvé que l’oxygène ne présente aucun effet direct sur l’activité azoréductase.
-
Effet du glucose : Le glucose a été décrit comme un substrat préféré pour la biodécoloration dans des conditions strictement anaérobie, alors qu’il ne l’est pas pour la biodégradation des colorants dans des conditions limitées en oxygène par les bactéries aérobie, même si, dans ce cas, sa présence améliore la cinétique de décoloration. En effet, la décoloration de jaune mordant 3 par Sphingomonas xenophoga souche BN6 est considérablement augmentée par la présence de glucose.
Cependant, à des concentrations seuils, le glucose constitue un facteur limitant de la décoloration. Il a été montré qu’une concentration de 5 g•L-1 de glucose est inhibitrice de la décoloration de certains diazoïques par P. luteola.
Selon Chenet al. (2003) et Pandeyet al. (2007), l’effet négatif du glucose sur la décoloration anoxique peut être attribué soit à la chute de pH due à la formation d’acide, soit à une répression catabolique. Changet al. (2001) ont expliqué ce phénomène par la répression catabolique exercée par le glucose sur l’expression de certains gènes. En effet, le glucose inhibe la transcription des gènes AMP cycliques-dépendants (White, 1995) dont font partie les gènes codants pour les azoréductases (Chang et al., 2001).
-
Effet de la source d’azote : La régénération du NADH, cofacteur de l’azoréduction, nécessite une source d’azote comme de l’extrait de levure, de la peptone, du tryptone, du KNO3, (NH4)2SO4, NH4Cl, etc.
Yuet al. (2001) ont testé l’influence de différentes sources d’azote sur la décoloration de l’acide violet 7 par Pseudomonas GM3. La peptone s’est révélée comme la meilleure source d’azote pour la régénération de NADH, alors que le KNO3 limite la cinétique de décoloration et inhibe complètement celle-ci à des concentrations élevées.
L’effet inhibiteur du KNO3 sur la décoloration dans des conditions anaérobie ou limitées en oxygène a été expliqué par le fait que l’ion nitrate (NO3-), et sa forme réduite le nitrite (NO2-), sont utilisés par la souche bactérienne comme des accepteurs d’électrons (Wuhrmannet al., 1980). Ces derniers entrent en compétition avec l’azoréduction comme accepteurs finaux des électrons (Yuet al., 2001; Zissi et Lyberatos, 1996).
Cependant, l’effet bénéfique de la peptone sur la biodégradation des colorants azoïques a été étudié et confirmé par d’autres chercheurs (Changet al., 2001; Hu, 1994).
Effet de la température et du pH : Généralement l’activité décolorante la plus importante est obtenue pour la température et le pH optima de la croissance de la bactérie (Changet al., 2001; Wuhrmann et al., 1980; Yuet al., 2001).
-
Effet de la structure chimique des colorants azoïques : La dégradation des colorants azoïques ne dépend pas que de l’activité intracellulaire de l’azoréductase, mais aussi du transfert de ces colorants à travers la membrane plasmique. En effet, la perméabilité cellulaire aux colorants dépend fortement de leur structure chimique et des ramifications qu’ils peuvent porter (Wuhrmann et al., 1980).
Mechsner et Wuhrmann (1982) ont montré l’importance du transport des colorants azoïques sulfonés sur la cinétique apparente de décoloration. Ces auteurs ont comparé la biodégradation de colorants azoïques par des cellules intactes à celle obtenue par des cellules perméabilisées de Bacillus cereus. Ils ont notamment observé que la décoloration était plus rapide avec les cellules perméabilisées qu’avec les cellules natives.
3.3.5.2 Dégradation des colorants azoïques par la succession anaérobie/aérobie
La dégradation des colorants azoïques par des bactéries dans des conditions limitées ou dépourvues en oxygène présente l’inconvénient d’accumuler des azo-produits (amines) qui sont la plupart du temps très toxiques, voire carcinogènes (Chunget al., 1981; De Franceet al., 1986; Junget al., 1992; Yahagiet al., 1975).
L’oxydation des ces amines aromatiques par les bactéries nécessite la présence d’oxygène, qui est, comme on l’a vu, un facteur limitant de l’azoréduction. Les chercheurs ont tenté de résoudre ce problème en combinant la biodégradation anaérobie et aérobie de manière à minéraliser complètement les colorants azoïques. Deux méthodes ont été alors étudiées :
-
Un système de réacteur séquentiel anaérobie/aérobie : Dans ce système, les colorants azoïques sont exposés en présence de la bactérie étudiée, dans un premier temps à des conditions anaérobie, puis dans un deuxième temps, à des conditions aérobie (Basibuyuk et Forster, 1997; O’Neillet al., 1999).
Cependant, les colorants azoïques n’ont jamais été minéralisés de façon satisfaisante dans la majorité des cas étudiés. Les amines aromatiques formées lors de la première étape de la biodégradation (azoréduction) s’accumulent dans le milieu extracellulaire et résistent à la bio-oxydation. La dégradation des amines aromatiques, en particulier celle des amines sulfoniques, est limitée, voire inhibée par leur faible transfert à travers la membrane cellulaire (Tanet al., 2005).
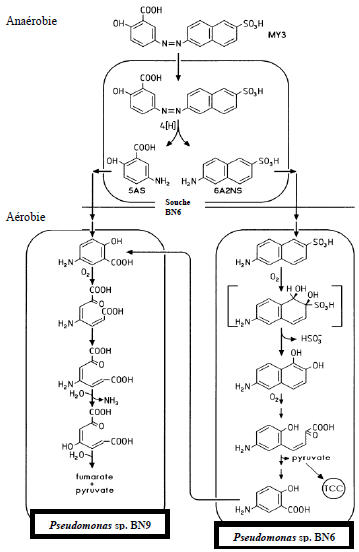
Parmi de nombreuses études, signalons la seule réalisée avec un consortium formé de deux Pseudomonas (Figure 5), qui fait l’exception et qui a abouti à la minéralisation totale du Jaune mordant 3 (Hauget al., 1991).
Figure 5
Minéralisation du colorant azoïque jaune mordant 3 (MY3) à l’aide d’un système de réacteur séquentiel anaérobie/aérobie (Haug et al., 1991).
Figure 5. Mineralization of azo dye mordant yellow 3 (MY3) by the sequential anaerobic/aerobic reactor (Haug et al., 1991).
Un système de réacteur anaérobie/aérobie intégré : Dans ce système, des microorganismes anaérobie et aérobie peuvent collaborer pour une minéralisation des colorants azoïques (Libraet al., 2004; Van der Zee et Villaverde, 2005). Un des problèmes majeurs est le déséquilibre entre le cosubstrat et l’oxygène; ceci est susceptible d’inhiber, notamment dans l’étape aérobie, la dégradation des amines aromatiques.
3.3.5.3 Dégradation des colorants azoïques par des bactéries dans des conditions oxygénées
Si la biodégradation des colorants azoïques dans des conditions anaérobie a été très largement étudiée, la bio-décoloration en culture aérée (bien oxygénée) reste encore peu étudiée et limitée à certaines espèces de bactéries. Ces dernières échappent aux conditions générales d’azoréduction en anaérobiose (Tableau 6).
Tableau 6
Principaux travaux portant sur la biodégradation des colorants azoïques dans des conditions oxygénées.
Table 6. Main works concerning the biodegradation of azo dyes in oxygenated conditions.

Certaines de ces bactéries, telles que Enterobacter agglomerans (Moutaouakkilet al., 2003; Moutaouakkilet al., 2004) et Pseudomonas fluorescens NCIM 2100 (Pandey et Upadhyay, 2006) se limitent à l’azoréduction, première étape de la biodégradation.
D’autres sont capables d’utiliser les amines aromatiques formées comme sources uniques de carbone, aboutissant ainsi à une dégradation complète ou « minéralisation » des colorants azoïques. Ces bactéries dégradent les molécules colorantes par la succession de deux étapes enzymatiques : une azoréduction, non sensible à l’oxygène, suivie d’une métabolisation à oxygène dépendant (Blümelet al., 1998; Wong et Yuen, 1998). Les Pseudomonas sont les plus citées et ont montré une grande aptitude à la conversion aérobie de colorants azoïques.
Ces bactéries expriment des azoréductases non sensibles à la présence de l’oxygène. En effet, l’orange I azoréductase [NADH(P)H:1-(4-sulfophenylazo)-4-naphtol oxydoréductase] et l’orange II azoréductase [NADH(P)H:1-(4-sulfophenylazo)-2-naphtol oxydoréductase], azoréductases non sensibles à l’effet de l’oxygène, ont été purifiées et caractérisées chez la souche Pseudomonas KF46 (Kullaet al., 1983; Zimmermannet al., 1982). En 2002, une équipe allemande a identifié la séquence des acides aminés de l’orange II azoréductase de Pseudomonas KF46 (Blumelet al., 2002) dont la structure était complètement différente de celle de Bacillus sp. OY12. Il a été montré par ailleurs que l’activité de cette azoréductase dépend fortement de la structure des colorants azoïques. En effet, Zimmermannet al. (1982) ont établi une corrélation entre l’activité de l’azoréductase et la structure chimique du colorant et ont souligné le rôle de certains groupements électrophiles dans la résistance du colorant à l’attaque enzymatique. Ils ont étudié, notamment, les effets des colorants, d’une part en tant qu’inducteur, et, d’autre part, en tant que substrat sur respectivement l’expression et l’activité de l’azoréductase.
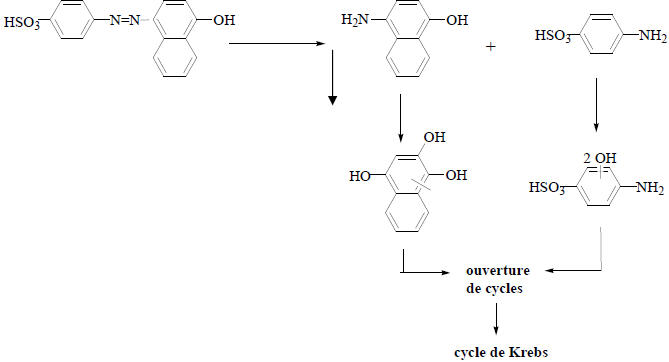
Après l’azoréduction vient l’oxydation : les amines aromatiques issues de l’azoréduction sont directement prises en charge par un système de cascade enzymatique impliquant des oxygénases et aboutissant à leur dégradation totale (minéralisation). Des amines aromatiques sulfoniques, connues comme étant très résistantes à l’attaque bactérienne à cause de leur faible transfert à travers la membrane cellulaire (Feigel et Knackmuss, 1993; Tan et al., 2005), ont été facilement biodégradables par des bactéries de genre Pseudomonas (Figure 6).
Figure 6
Mécanisme de dégradation de l’acide orange 20 par Pseudomonas K24 (Kulla et al., 1983).
Figure 6. Degradation mechanism of acid orange 20 by Pseudomonas K24 (Kulla et al., 1983).
3.3.6 Résumé de procédés biologiques et analyse critique
La dégradation des colorants synthétiques a été décrite au moyen de microorganismes tels que les champignons, les algues, les actinomycètes, les levures et a montré un certain nombre d’avantages parmi lesquels on peut citer :
L’efficacité dans la dégradation des colorants azoïques sulfonés connus comme étant très résistants à l’attaque bactérienne (Banatet al., 1996);
La dégradation extracellulaire des colorants qui élimine la difficulté liée à la filtration de la molécule colorante par la membrane cellulaire.
Le peu d’influence sur ces microorganismes des paramètres physico-chimiques connus comme facteurs limitants dans le cas des bactéries.
Mais elle est aussi caractérisée par un certain nombre d’inconvénients :
Le cycle de croissance très long des champignons et des algues limite leurs performances à décolorer;
Ces microorganismes sont difficiles à manipuler dans des assez grands volumes.
La dégradation extracellulaire rejette directement dans le milieu les produits qui en dérivent et qui peuvent être très toxiques pour l’environnement;
Dans certains cas, la décoloration par les champignons et les actinomycètes est due à une adsorption membranaire du colorant et non à une biodégradation (Zhou et Zimmermann, 1993), ce qui déplace le problème puisque le colorant reste intact et la toxicité demeure.
Ce constat a beaucoup contribué à l’orientation de beaucoup de chercheurs vers le traitement des colorants synthétiques par les bactéries qui soit plus efficace et qui présente moins d’inconvénients. Les bactéries se sont révélées très efficaces sur les colorants, en particulier les azoïques. Toutefois, certaines bactéries se limitent à la première étape de la dégradation « azoréduction » donnant naissance à des amines aromatiques généralement plus toxiques que la molécule mère alors que d’autres sont capables d’emmener la biodégradation jusqu’à la fin « minéralisation ». Pour cela, plusieurs procédés ont été développés afin d’améliorer la biodégradabilité de colorants, en optimisant les conditions de culture en oxygène, température, pH et la richesse en sources de carbone (glucose) et d’azote (extrait de levure, peptone, tryptone etc.).
4. Conclusion
Les colorants de synthèse sont de plus en plus utilisés dans les industries en raison de leur facilité de synthèse, de leur rapidité de production et de leur variété de couleurs si on les compare aux colorants naturels. La diversité structurale des colorants synthétiques est due à la fois à la diversité des groupements chromophoriques qui les composent (groupements azoïque, anthraquinone, triarylmethane et phtalocyanine) et à la diversité des technologies d’application (coloration réactive, directe, dispersée ou de cuve). Le secteur des colorants constitue une véritable industrie qui est impliquée dans un grand éventail de domaines : industrie textile, imprimerie, industrie pharmaceutique, cosmétique et agro-alimentaire. L’industrie des colorants constitue aujourd’hui un secteur important de la chimie : la production mondiale est estimée à plus de 800 000 tonnes par an dont 140 000 sont perdues dans les effluents au cours des différentes étapes d’application et de confection (Cooper, 1995; Zollinger, 1987).
Parmi les nombreuses familles de colorants synthétiques, les colorants azoïques sont les plus largement utilisés (60 à 70 %). Ces colorants constituent un groupe de composés caractérisés par une ou plusieurs liaisons azoïques (R1-N = N-R2) en association avec un ou plusieurs groupements aromatiques, ce qui les rend très stables et relativement peu biodégradables. Beaucoup d’études ont montré, d’autre part, des effets toxiques et/ou carcinogènes de colorants azoïques (Medvedevet al., 1988; Miller et Miller, 1961; Umbuzeiroet al., 2005; Yahagiet al., 1975), ce qui contraint à traiter les effluents contenant ces colorants avant de les rejeter dans le milieu naturel.
Les traitements physico-chimiques traditionnels (adsorption, coagulation/floculation, précipitation etc.) sont couramment utilisés pour la dépollution des effluents industriels; bien qu’elles soient rapides, ces méthodes s’avèrent généralement peu efficaces en regard des normes exigées sur les rejets. Elles sont très coûteuses et chargent les rejets finaux en nombreux produits chimiques. Il apparaît donc intéressant de mettre au point des traitements alternatifs, notamment par voie biologique, qui ont l'avantage d'être moins coûteux, moins polluants et plus efficaces car plus spécifiques (Moutaouakilet al., 2004).
Les colorants azoïques sont transformés par certaines variétés de microorganismes, incluant des bactéries (aérobies et anaérobies) et des champignons. Ces derniers ont généralement des croissances et des cinétiques de décoloration lentes, ce qui limite leurs performances vis-à-vis de ces colorants. Par contre, il a été rapporté (Basibuyuk et Forester, 1997; Nachiyar et Rajakumar, 2004) que la dégradation complète (minéralisation) des colorants azoïques par les bactéries nécessite la succession de deux étapes : une azoréduction (anaérobie) suivie d’oxydations aérobies des amines aromatiques formées lors de l’étape précédente. Certaines bactéries se limitent cependant à la première étape de la biodégradation ou « azoréduction » donnant naissance à des amines aromatiques qui s’accumulent dans le milieu de culture. Des colorants qui sont parfois non toxiques à l'origine peuvent ainsi évoluer par ce processus en métabolites toxiques : mutagènes, voire cancérigènes (Rafiiet al., 1997). Néanmoins, les Pseudomonas sont des bactéries très actives sur ce type de molécules et expriment fortement des azoréductases (Changet al., 2001; Hu, 2001). Parmi ce genre bactérien, certaines souches sont capables de minéraliser complètement ce type de colorants (Nachiyar et Rajakumar, 2004).
Parties annexes
Références bibliographiques
- Abraham T.E., R.C. Senan, T.S. Shaffiqu, J.J. Roy, J.J., T.P. Poulouse et P.P. Thomas (2003). Bioremediation of textile azo dyes by an aerobic bacterial consortium using a rotating biological contactor. Biotechnol. Progr., 19, 1372-1376.
- Adedayo O., S. Javadpour, C. Taylor, W.A. Anderson et M. Moo-Young (2004). Decolorization and detoxification of methyl red by aerobic bacteria from a wastewater treatment plant. World J. Microbiol. Biotechnol., 20, 545–550.
- Alvares A.B.C., C. Dlaper et S.A. Parsons (2001). Partial oxidation by ozone to remove recalcitrance from wastewaters – a review. Environ. Technol., 22, 409-427.
- Alves de Lima R.O., A.P. Bazo, D.M. Favero Salvadori, C.M. Rech, d. De Palma Oliveira et G. de A. Umbuzeiro (2007). Mutagenic and carcinogenic potential of a textile azo dye processing plant effluent that impacts a drinking water source. Mutation Res., 626, 53-60.
- ANSELME C. et E.P. JACOBS (1996). Water treatment membrane processes. McGraw Hill Mallevialle, New York, NY, USA, pp. 401-1087.
- Azbar N., T. Yonar, T. et K. Kestioglu (2004). Comparison of various oxidation processes and chemical treatment methods for COD and color removal from a polyester and acetate fiber dyeing effluent. Chemosphere 55, 1, 35-43.
- Balan D.S.L. et R.T.R. Monteiro (2001). Decolorization of textile indigo dye by ligninolytic fungi. J. Biotechnol., 89, 141–145.
- Ball A.S., W.B. Betts et A.J. McCarthy (1989). Degradation of lignin-related compounds by Actinomycetes. Appl. Environ. Microbiol., 55, 1642-1644.
- Banat I.M., P. Nigam, D. Singh et R. Marchant (1996). Microbial decolorization of textile-dye-containing effluents: A review. Bioresour. Technol., 58, 217-227.
- Banerjee A., D. Nabasree et B. De (2005). In vitro study of antioxidant activity of Syzygium cumini fruit. Food Chem., 90, 727-733.
- Basibuyuk M. et C.F. Forester (1997). The use of sequential anaerobic/aerobic processes for the biotreatment of a simulated dyeing wastewater. Environ. Technol., 18, 843-848.
- Bauer C., P. Jacques et A. Kalt (2001). Photooxidation of an azo dye induced by visible light incident on the surface of TiO2. J. Photochem. Photobiol. A Chem., 140, 87-92.
- Ben Mansour H., D. Corroler, D. Barillier, K. Ghedira, L. Chekir et R. Mosrati (2007). Evaluation of genotoxicity and pro-oxidant effect of the azo dyes: Acids yellow 17, violet 7 and orange 52, and their biodegradation products by Pseudomonas putida mt-2. Food Chem. Toxicol., 45, 1670-1677.
- Ben Mansour H., R. Mosrati, D. Corroler, D. Bariller, K. Ghedira, D. Bariller et L. Chekir-Ghedira (2009a) In vitro study of DNA damage induced by acid orange 52 and its biodegradation derivatives. Environ. Toxicol. Chem., 28, 489-495.
- Ben Mansour H., R. Mosrati, D. Corroler, K. Ghedira, D. Bariller et L. Chekir (2009b). Genotoxic and anti-butyrylcholinesterasic activities of acid violet 7 and its biodegradation products. Drug Chem. Toxicol., 32, 230-237.
- Ben Mansour H., D. Bariller, D. Correler, K. Ghedira, L. Chekir et R. Mosrati (2009c). In vitro mutagenicity of acid violet 7 and its degradation products by Pseudomonas putida mt-2: Correlation with chemical structure. Environ. Toxicol. Pharmacol., 27, 231-236.
- Ben Mansour H., R. Mosrati, D. Corroler, K. Ghedira, D. Bariller et L. Chekir (2009d). Genotoxic and anti-butyrylcholinesterasic activities of acid violet 7 and its biodegradation products. Drug Chem. Toxicol., 32, 230-237.
- Ben Mansour H., D. Corroler, D. Bariller, K. Ghedira, L. Chekir-Ghedira et R. Mosrati (2009e). Influence of the chemical structure on the biodegradability of acids yellow 17, violet 7 and orange 52 by Pseudomonas putida. Ann. Microbiol., 59, 1-7.
- Ben Mansour H., R. Mosrati, D. Corroler, K. Ghedira, D. Bariller et L. Chekir-Ghedira (2009f) Mutagenicity and genotoxicity of acid yellow 17 and its biodegradation products. Drug Chem. Toxicol., 32, 222-229
- Ben Mansour H., R. Mosrati, D. Corroler, K. Ghedira, D. Bariller et L. Chekir-Ghedira (2010). Acid violet 7 and its biodegradation products induce chromosome aberration, lipid peroxidation and cholinesterase inhibition in mouse bone morrow. Environ. Sci. Pollut. Res., 17, 1371-1778.
- Benitez F.J., J.L. Acero, F.J. Real et A.I. Leal (2001). The role of hydroxyl radicals for the decomposition of p-hydroxy phenylacateic acid in aqueous solutions. Water Res., 35, 1338-1343.
- Bessekhouad Y., D. Roberto et J.V. Weber (2003). Synthesis of photocatalytic TiO2 nanoparticles: optimization of the preparation conditions. J. Photochem. Photobiol. A Chem., 157, 47-53.
- Blumel S., H.J. Knackmuss et A. Stolz (2002). Molecular cloning and characterizatiion of the gene coding for the aerobic azoreductase from Xenophillus azovorans KF44F. Appl. Environ. Microbiol., 68, 3948–3955.
- Blümel S., C. Mattias, L. Martina, S. Ansreas et K. Hans-Joachim (1998). Isolation of a bacterial strain with the ability to utilize the sulfonated azo compound 4-carboxy sulfoazobenzene as the sole source of carbon and energy. Appl. Environ. Microbiol., 64, 2315–2317.
- Brohm K. et E. Frohwein (1937). Nachweis von durch Säueringentfarbten künstlichen Eigelbfrabstoffen in Milchspeiseeis. Zbl. Lebensmitt. Forsch., 73, 30.
- Brown M.A. et S.C. Devito (1993). Predicting azo dye toxicity. Crit. Rev. Environ. Sci. Technol., 12, 405-414.
- Burke NS. et D.L. Crawford (1998). Use of azo dye ligand chromatography for the partial purification of a novel extracellular peroxidase from Streptomyces viridosporus T7A. Appl. Microbiol. Biotechnol., 49, 523-30.
- Calabro V., G. Pantano, R. Kang, R. Molinari et E. Drioli (1990). Experimental study on integrated membrane processes in the treatment of solutions simulating textile effluents. Energy and exergy analysis. Desalination, 78, 257-277.
- CAPON M., V. COURILLEU et C. VALTTE (1999). Chimie des couleurs et des odeurs, culture et technique. Nantes ISBN 2-9502444-2-4
- Chang J.S., Chou, C., Y.C. Lin, P.J. Lin, J.Y. Ho et T.L. Hu (2001). Kinetic characteristic of bacterial azo dyes decolorization by Pseudomonas luteola. Water Res., 35, 2841-2850.
- Chang J.S. et Y.C. Lin (2000). Fed-batch bioreactor strategies for microbial decolorization of azo dye using a Pseudomonas luteola strain. Biotechnol. Prog., 16, 979-985.
- Chen K.C., J.Y. Wu, D.J. Liu et S.C.J. Hwang (2003). Decolorisation of the textile dyes by newly isolated bacterial strains. J. Biotechnol., 101, 57-68.
- Chen H., S.L. Hopper et C.E. Cerniglia (2005). Biochemical and molecular characterization of an azoreductase from Staphylococcus aureus, a tetrameric NADPH-dependent flavoprotein. Microbiology 151, 1433–1441.
- Chen B.Y. (2006). Toxicity assessment of aromatic amines to Pseudomonas luteola: chemostat pulse technique and dose-response analysis. Proc. Biochem., 41, 1529–1538.
- Chen H., R.F. Wang et C.E. Cerniglia (2004). Molecular cloning, overexpression, purification, and characterization of an aerobic FMN-dependent azoreductase from Enterococcus faecalis. Protein Expr. Purif., 34, 302–310.
- Chivukula M. et V. Renganathan (1995). Phenolic azo dye oxidation by laccase from Pyricularia oryzae. Appl. Environ. Microbiol., 61, 4374-4377.
- Christie R. (2001). Colour chemistry. The Royal Society of Chemistry, Cambridge, United Kingdom.
- Chung KT., E.G. Fulk et A.Y. Andrews (1981). Mutagenicity testing of some commonly used dyes. Appl. Environ. Microbiol., 42, 641-648.
- Chung K.T., G.E. Fulk et M. Egan (1978). Reduction of azo dyes by intestinal anaerobes. Appl. Environ. Microbiol., 35, 5588-5620.
- Chung K.T., S.E.J. Stevens et C.E. Cerniglia (1992). The reduction of azo dyes by the intestinal microflora. Critical Rev. Microbiol., 18, 175–197.
- Combes R.D. et R.B. Haveland-Smith (1982). A review of the genotoxicity of food, drug, and cosmetic colour and other azo, triphenylmethane and xanthene dyes. Mutation Res. Rev. Genetic Toxicol., 198, 101-243.
- COOPER P (1995). Color in dye house effluent. Society of dyes and colourists (Éditeur), Bradford, West Yorkshire, Grande-Bretagne, 197 p.
- Crips C., A. John, J.A. Bumpus et S.D. Aust (1990). Biodegradation of azo and heterocyclic dyes by Phanerochaete chrysosporium.Appl. Environ. Microbiol., 56, 1114-1118.
- De France B.F., M.H. Carter et P.D. Josephy (1986). Comparative metabolism and mutagenicity of azo and hydrazone dyes in the Ames test. Food Chem. Toxicol., 24, 165-196.
- De Heridia J.B., J. Torregrosa, R. Dominguez et J.A. Peres (2001). Kinetic model for phenolic compound oxidation by Fenton’s reagent. Chemopshere, 45, 85-90.
- DEPA (Danish Environmental Protection Agency) (2000). Survey of azo-colorants in Denmark, Toxicity and fate of azo dyes.
- Dos Santos A., F.J. Cervantes et J.V. van Lier, (2007). Review paper on current technologies for decolourisation of textile wastewaters: Perspectives for anaerobic biotechnology. Bioresour. Technol., 98, 2369-2385.
- Durán N., M.F.S. Teixeira, R. De Conti et E. Esposito (2002). Ecological-friendly pigments from fungi. Critical Rev. Food Sci. Nutr., 42, 53-66.
- Eichlerova I., L. Homolka, L. Lisa et F. Nerud (2005). Orange G and remazol brilliant blue R decolorization by white rot fungi Dichomitus squalens, Ischnoderma resinosum and Pleurotus calyptratus. Chemosphere, 60, 398–404.
- Fang H., H. Wenrong et L. Yuezhong (2004). Biodegradation mechanisms and kinetics of azo dye 4BS by a microbial consortium. Chemosphere, 57, 293–301.
- Feigel B.J. et H.J. Knackmuss (1993). Syntropic interactions during degradation of 4 aminobenzenesulfonic acid by a two species bacterial culture. Arch. Microbiol., 159, 124–130.
- Fenton H.J.H. (1894). Oxidation of tartaric acid in the presence of ion. J. Chem. Soc., 65, 899-910.
- Fujian X., C. Hongzhang et L. Zuohu (2001). Solid–state production of lignin peroxidase (LiP) and manganese peroxidase (MnP) by Phanerochaete chrysosporium using steam-exploded straw as substrate. Bioresour. Technol., 80, 149-151.
- Galindo C. (1998). Dégradation de colorants par la méthode d’oxydation avancée UV/H2O2. Thèse de doctorat, n° 98 MULH 0520, Université de Mulhouse, France.
- Ganesh R., g.d. Boardman ET D. Michelson (1994). Fate of azo dyes in sludges. Water Res., 28, 1367-1376.
- GANESH R. (1992) Fate of azo dye in sludges. Thèse de doctorat, Chimie, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 193 p.
- Ghorpade A.K. et H.T. Spencer (1993). Azo dyes metabolism by Pseudomonas putida. Dans : 48th Purdue International Waste Conference Proceedings, Lewis Publishers, Chelsea, Michigam, pp. 699-714.
- Glenn J.K. et M.H. Gold (1983). Decolorization of several polymeric dyes by the lignin- degrading basidiomycete Phanerochaete chrysosporium. Appl. Environ. Microbiol., 45, 1741-1747.
- Golka K., S. Kopps et Z.W. Myslak (2004). Carcinogenicity of azo colorants: influence of solubility and bioavaibility. Toxicol. Lett., 151, 203-210.
- Guivrach E. et M.A. Oturan (2004). Le problème de la contamination des eaux par les colorants synthétiques : comment les détruire? Actualité chim., 277/238, 65-68.
- Haber F. et J. Weiss (1934). The catalytic decomposition of hydrogen peroxide by iron salts. Proc. Nat. Acad. Sci., 134, 332-351.
- Hao O.J., H. Kim et P.C. Chiang (2000). Decolorization of wastewater. Crit. Rev. Environ. Sci. Technol., 30, 449-505.
- Harazono, K. et K. Nakamura (2005). Decolorization of mixtures of different reactive textile dyes by the white-rot basidiomycete Phanerochaete sordida and inhibitory effect of polyvinyl alcohol. Chemosphere, 59, 63-68.
- Haug, W., A. Schmidt, B. Nortemann, D.C. Hempel, A. Stolz et H.J. Knackmuss (1991). Mineralization of the sulphonated azo dye mordant yellow 3 by a 6-aminonaphthatene- 2-sulphonate-degrading bacterium consortium. Appl. Environ.l Microbiol., 57, 3144-3149.
- Hernandez, R., M. Zappi, J. Colucci et R. Jones (2002). Comparing the performance of various advanced oxidation processes for treatment of acetone contaminated water. J. Hazard Mater., 92, 33-50.
- Hong, A., M.E. Zappi, C.H. Kuo et D.O. Hill (1996). Modelling the kinetics of illuminated and dark advanced oxidation processes. ASCE J. Environ. Eng., 122, 1, 58-62.
- Horitsu, H., M. Takada, E. ldaka, M. Tomoyeda et T. Ogawa (1977). Degradation of p-aminoazobenzene by Bacillus subtilis. Eur. J. Appl. Microbiol., 4, 217-224.
- HSUEH C.-C. et B.-Y. Chen (2007). Comparative study on reaction selectivity of azo dye decolorization by Pseudomonas luteola. J. Hazard. Mater., 141, 842-849.
- Hu, T.L. (1994). Decolourization of reactive azo dye by transformation with Pseudomonas luteola. Bioresour. Technol., 49, 47–51.
- Hu, T.L. (2001). Kinetics of azoreductase and assessment of toxicity of metabolic products from azo dyes by Pseudomonas luteola. Water Sci. Technol., 43, 2, 261–269.
- IARC (1982). World Health Organization, International Agency for research on Cancer. Dans : Monographs on the evaluation of the carcinogenic risk of chemicals to human. "Some industrial chemicals and dyestuffs", Lyon, France, 29 p.
- Idaka E. et Y. Ogawa (1978). Degradation of azo compounds by Aeromonas hydrophila var. 2413. Dyers Colour., 94, 91-94.
- Ince N.H. et G. Tezcanli (2001). Reactive dyestuff degradation by combined sonolysis and ozonation. Dyes Pigments, 49, 145-153.
- Jinqi L. et L. Houtian (1992). Degradation of azo dyes by algae. Environ. Pollut., 75, 273-278.
- Jung R., D. Steinle et R. Anliker (1992). A compilation of genotoxicity and carcinogenicity ata of aromatic aminosulfonic acids. Food Chem. Toxicol., 30, 635-660.
- Kalme S.D., G.K. Parshetti, S.U. Jadhav et S.P. Govindwar (2007). Biodegradation of benzidine based dye Direct Blue-6 by Pseudomonas desmolyticum NCIM 2112. Bioresour. Technol., 98, 1405–1410.
- Kaushik G., M. Gopal et I.S. Thakur (2010). Evaluation of performance and community dynamics of microorganisms during treatment of distillery spent wash in a three stage bioreactor. Bioresour. Technol., 101, 4296-4305.
- Kondo M.M., M.A.S.V. Arcos, T. Marco, T. et M.T. Grassi (2002). Dissolved organic carbon determination using FIA and photo-fenton reaction. Brazil. Arch. Biol. Technol., 45, 81-87.
- Kulla H.G., F. Klausener, U. Meyer, B. Lüdeke et T. Leisinger (1983). Interference of aromatic sulfo groups in the microbial degradation of the azo dyes orange I and orange II. Arch. Microbiol., 135, 1-7.
- Kuo W.G. (1992). Decolorization dye wastewater with feteon’s reagent. Water Res.. 26, 881-886.
- Libra J.A., M. Borchert, L. Vigelahn et T. Thomas Storm (2004). Two stage biological treatment of a diazo reactive textile dye and the fate of the dye metabolites. Chemosphere, 56, 167-180.
- Lide D.R. (1999). Hanbook of chemistry and physics. Solubility of selected gases in water. 79e Ed., Cleveland (OH), Chemical Rubber Co.
- Lin C.C. et Y.T. Lai (2006). Adsorption and recovery of lead (II) from aqueous solutions by immobilized Pseudomonas aeruginosa PU21 beads. J. Hazard. Mater., 137, 99-105.
- Lin C.N., H.L. Chen et M.H. Yen (2008). Flavonoids with DNA strand-scission activity from Rhus javanica var. roxburghiana. Fitoterapia, 79, 32–36.
- Maier J., A. Kandelbauer, A. Erlacher, A. Cavaco-Paulo et G.M. Gübitz (2004). A new alkali-thermostable azoreductase from Bacillus sp. strain SF. Appl. Environ. Microbiol., 70, 837–844.
- MANAHAN S.E (1994). Environmental chemistry. Lewis publishing, 6e édition, Atlanta, GA, USA.
- Manning B.W., C.E. Cernigli et T.E. Federle (1985). Metabolism of the benzidine-based azo dye direct black 38 by human intestinal microbiota. Appl. Environ. Microbiol., 50, 10-15.
- Mechsner K. et K. Wuhrmann (1982). Cell permeability as a rate limiting factor in the microbial reduction of sulfonated azo dyes. Eur. J. Appl. Microbiol. Biotechnol., 15, 123-126.
- Medvedev Z.A., H.M. Crowne et M.N. Medvedeva (1988). Age related variations of hepatocarcinogenic effect of azo dye (3’-MDAB) as linked to the level of hepatocyte polyploidization. Mech. Ageing Develop., 46, 159-174.
- Miller J.A. et E.C. Miller (1961). The carcinogenicity of 3-methoxi-4-aminoazo-benzen and its N-methyl derivatives for extrahepatic tissues of the rat. Cancer Res., 21, 1068-1074.
- Mills C., R.J. Bull et K.P. Cantor (1998). Risques pour la santé liés à la consommation de sous-produits de la chloration de l'eau potable : rapport d'un groupe d'experts. Maladie Chron. Canada, 19, 3.
- Moutaouakkil M., Y. Zeroual, F.Z. Dzayri, M. Talbi, K. Lee et M. Blghen (2003). Purification and partial characterization of azoreductase from Enterobacter agglomerans. Arch. Biochem. Biophys., 413, 139–146.
- Moutaouakkil M., Y. Zeroual, F.Z. Dzayri, M. Talbi, K. Lee et M. Blghen (2004). Decolorization of azo dyes with Enterobacter agglomerans immobilized in different supports by using fluidized bed bioreactor. Current Microbiol., 48, 124–129.
- Nachiyar C.V. et G.S. Rajakumar (2004). Mechanism of Navitan Fast Blue S5R degradation by Pseudomonas aeroginosa. Chemosphere, 57, 165–169.
- Neamtu M., A. Yediler, I. Siminicanu, M. Macoveanu et A. Kettrup (2004). Decolorization of disperse red 354 azo dye in water by several oxidation processes – a comparative study. Dyes Pigment, 60, 61-68.
- Nigam P., I.M. Banat, D. Singh et R. Marchant (1996). Microbial process for the decolorization of textile effluent containing azo, diazo and reactive dyes. Proc. Biochem., 31, 435-442.
- Nigam, P. et R. Marchant (1995). Selection of a substratum for composing biofilm system of a textile-effluent decolorizing bacteria. Biotechnol. Lett., 17, 993-996.
- O’Neill, C., F.R. Hawkes, D.L. Hawkes, N.D. Lourenco, H.M. Pinheiro et W. Delee (1999). Colour in textile effluents – sources, measurement, discharge consents and simulation: a review. J. Chem. Technol. Biotechnol., 74, 10009–10018.
- Pagga, U. et D. Brown (1986). The degradation of dyestuffs part II: behaviour of dyestuffs in aerobic biodegradation tests. Chemosphere, 15, 479-491.
- Pandey, A, P. Singh et L. Iyengar (2007). Bacterial decolorization and degradation of azo dyes. Int. Biodeter. Biodegrad., 59, 73-84.
- Pandey, B.V. et R.S. Upadhyay (2006). Spectroscopic characterization and identification of Pseudomonas fluorescens mediated metabolic products of Acid Yellow-9. Microbiol. Res., 161, 311-315.
- Pasti, M.B., A.L. Pometto et M.P. Nuti (1990). Crawford DL.lLignin-solubilizing ability of actinomycetes isolated from termite (Termitidae) gut. Appl. Environ. Microbiol., 56, 2213-2218.
- Paszczynski A., M.B. Pasti-Grigsby, S. Goszczynski, R.L. Crawford et D.L. Crawford (1992). Mineralization of sulfonated azo dyes and sulfanilic acid by Phanerochaete chrysosporium and Streptomyces chromofuscust. Appl. Environ. Microbiol., 58, 3598-3604.
- Percy A.J., N. Moore et J.K. Chipman (1989). Formation of nuclear anomalies in rat intestine by benzidine and its biliary metabolites. Toxicology, 57, 217-223.
- Quillardet P. et M. Hofnung (1993). The SOS chromotest: a review. Mutation Res./Rev. Gen. Toxicol., 297, 235-279.
- Rafii F. et C.E. Cerniglia (1993). Comparison of the azoreductase and nitroreductase from Clostridium perfringens. Appl. Environ. Microbiol., 59, 1731-1734.
- Rafii F., J.D. Hall et C.E. Cernigalia (1997). Mutagenicity of azo dyes used in foods, drugs and cosmetics before and after reduction by Clostridium species from the human intestinal tract. Food Chem. Toxicol., 35, 897-901.
- Raghavacharya C. (1997). Colour removal from industrial effluents – a comparative review of available technologies. Chem. Eng. World, 32, 53-54.
- RAMALHO, DCF (2005). Degradation of dyes with microorganisms studies with Ascomycete yeasts., Université de Minho, Portugal, 14 p.
- Ramalho P.A., H. Scholze, M.H. Cardoso, M.T. Ramalho et A.M. Oliveira-Campos (2002.) Improved conditions for the aerobic reductive decolourisation of azo dyes by Candida zeylanoides. Enzyme Microb. Technol., 31, 848-854.
- Rau J., H.J. Knackmuss et A. Stolz (2002). Effects of different quinoid redox mediators on the anaerobic reduction of azo dyes by bacteria. Environ. Sci. Technol., 36, 1497–1504.
- Rehn L. (1895). Blasengeschwulste bei Fuschin arbeiten. Arch. Klin Chir., 50, 588.
- Ren S., J. Guo et G. Zeng (2006). Guoping sun decolorization of triphenylmethane, azo, and anthraquinone dyes by a newly isolated Aeromonas hydrophila strain. Appl. Microbiol. Biotechnol., 72, 1316–1321.
- Robinson T., G. McMullan, R. Marchant et P. Nigam (2001). Remediation of dyes in textile effluent: a critical review on current treatment technologies with a proposed alternative. Bioresour. Technol., 77, 247-255.
- Russ R., J. Rau et A. Stolz (2000). The function of cytoplasmic flavin reductases in the reduction of azo dyes by bacteria. Appl. Environ. Microbiol., 66, 1429–1434.
- Sandhu P. et J.K. Chipman (1990). Bacterial mutagenesis and hepatocyte unscheduled DNA synthesis induced by chrysoidine azo-dye components. Mutation Res., 240, 227-236.
- Santé Canada (1999). Chloration de l'eau, votre santé et vous.
- Seyewetz A. et P. Sisley (1896). Chimie des matières colorantes artificielles.Libraires de l’Académie de Médicine (Éditeur), Paris Masson, France.
- Shu H.Y. et C.R. Huang (1995). Degradation of commercial azo dyes in water using ozonation and UV enhanced ozonation process. Chemosphere, 31, 3813-3825.
- Slokar Y.M. et A.M. Le Marechal (1998). Methods of decoloration of textile wastwaters. Dyes Pigments, 37, 335-356.
- Solozhenko E.G., N.M. Soboleva et V.V. Goncharuk (1995). Decolorization of azo dye solutions by Fenton’s oxidation. Water Res., 29, 2206-2210.
- Sugiura W., T. Miyashita, T. Yokoyama et M. MOT00 Ara (1999). Isolation of azo-dye-degrading microorganisms and their application to white discharge printing of fabric. J. Biosci. Bioeng., 88, 577-581.
- Sugiura W., T. Yoda, T. Matsuba, Y. Tanaka et Y. Suzuki (2006). Expression and characterization of the genes encoding azoreductases from Bacillus subtilis and Geobacillus stearothermophilus. Biosci. Biotechnol. Biochem., 70, 1655-1665.
- Suzuki T., S. Timofei, L. Kurunczi, U. Dietze et G. Schurmann (2001). Correlation of aerobic biodegradability of sulfonated azo dyes with the chemical structure. Chemosphere, 45, 1-9.
- Swamy J. et J.A. Ramsay (1999). Effects of glucose and NH4+ concentrations on sequential dye decoloration by Trametes versicolor. Enz. Microb. Technol.,, 25, 278-284.
- Szpyrkowicz L., C. Juzzolino et S.N. Kaul (2001). A comparative study on oxidation of disperses dyes by electrochemical process, ozone, hydrochlorite and Fenton reagent. Water Res., 35, 2129-2136.
- Tan N.C., A. van Leeuwen, E.M. van Voorthuizen et P. Slenders (2005). Prenafeta-Boldu, F.X.; Temmink, H.; Lettinga, G.; Field, J. A. Fate and biodegradability of sulfonated aromatic amines. Biodegrad., 16, 527-537.
- Taylor J.S. et E.P. Jacobs (1996). Water treatment membrane processes. McGRAW HILL (Éditeur), New-York, NY, 238 p.
- Tsuda S., N. Matsusaka, H. Madarame, S. Ueno, N. Susa, K. Ishida, N. Kawamura, K. Sekihashi et Y.F. Sasaki (2000). The comet assay in eight mouse organs: result with 24 azo compounds. Mutation Res., 465, 11-26.
- Tzitzi M., D.V. Vayenas et G. Lyberatos (1994). Pretreatment of textile industry wastewaters with ozone. Water Sci. Technol., 28, 151-160.
- Umbuzeiro G.A., H. Freeman, S.H. Warren, F. Kummrow et L.D. Claxton (2005). Mutagenicity evaluation of the commercial product CI Disperse Blue 291 using different protocols of the Salmonella assay. Food Chem. Toxicol., 43, 49-56.
- Vander Bruggen B., L. Lejon et C. Vandecasteele (2003). Reuse, treatment and discharge of the concentrate of pressure-driven membrane processes. Environ. Sci. Technol., 37, 3733-3738.
- Van der Zee F.P. et S. Villaverde (2005). Combined anaerobic–aerobic treatment of azo dyes-A short review of bioreactor studies. Water Res., 39, 1425-1440.
- Vendevivere P.C., R. Bianchi et W. Verstraete (1998). Treatement and creuse from the textile wet-processing industry: review of emerging technologies. J. Chem. Technol. Biotechnol., 72, 289-302.
- Venkataraman K. (1901). The analytical chemistry of synthetic dyes. National Chemistry Laboratory, Poona, India ISBN 0-471-90575-5.
- Walker R. (1970). The metabolism of azo compounds: a review of the literature. Food Cosm. Toxicol., 8, 659-676.
- Wang J., B. Guo, X. Zhang, Z. Zhang, J. Han et J. Wu (2005). Sonocatalytic degradation of methyl orange in the presence of TiO2 catalysts and catalytic activity comparison of rutile and anatase. Ultrason. Sonochem., 12, 331–337.
- Welham A. (2000). The theory of dyeing (and the secret of life). J. Soc. Dyers Colour., 116, 140-143.
- White D. (1995). The physiology and biochemistry of Prokaryotes. Oxford University Press, New York, NY.
- Willmott N.J., J.T. Gutherie et G. Nelson (1998). The biotechnology approach to colour removal from textile effluent. J. Soc. Dyers Colour., 114, 38-41.
- Wong P.K. et P.Y. Yuen (1998). Decolorization and biodegradation of methyl red by Klebsiella pneumoniae RS-13. Water Res., 30, 1736-1744.
- Wuhrmann K., K.I. Mechsner et T. Kappeler (1980). Investigation on rate determining factors in the microbial reduction of azo dyes. Appl. Microbiol. Biotechnol., 9, 325-338.
- Yahagi Y., M. Degawa, Y. Seino, T. Matsushima, M. Nagao, T. Sugimura et Y. Hashimoto (1975). Mutagenicity of mutagenic azo dyes and their derivatives. Cancer Lett., 1, 91-96.
- Yatome C., T. Ogawa, H. Hayashi et T. Ogawa (1991). Microbial reduction of azo dyes by several strains. J. Environ. Sci. Health, A 26, 471-485.
- Yatome C., T. Ogawa, D. Koga et E. Idaka (1981). Biodegradability of azo and triphenyl methane dyes by Pseudomonas pseudomallei 13NA. J. Soc.Dyers Colour., 97, 166-169.
- Yatome C., T. Ogawa, K. Itoh, A. Sugiyama et E. Idaka (1987). Degradation of azo dyes by cell-free extracts from Aeromonas hydrophila var. 2413. J. Soc. Dyers Colour., 3, 389-395.
- Yu J., X. Wang et P.L. Yue (2001). Optimal decolorization and kenetic modeling of synthetic dyes by Pseudomonas strains. Water Res., 35, 3579-3586.
- Yuseuf R.O. et J.A. Sonibare (2004). Characterization of textile industries effluents in Kaduna, Nigeria and pollution implications. Global Nest Int. J., 6, 212-221
- Zhang F., A. Yeldiler, X. Liang et A. Kettrup (2004). Effects of dye additives on the ozonation process and oxidation by-products: a comparative study using hydroxylzed C.I. Reactive Red 120. Dyes Pigments, 60, 1-7.
- Zhenwang L., C. Zhenlu et L. Jianyan (2000). The PT dye molecular structure and its chromophoric luminescences mechanism. Dans : 15th World Conference on Non-Destructive Testing, 15-21 octobre 2000, Rome, Italie.
- Zhou W. et W. Zimmermann (1993). Decolorization of industrial effluents containing reactive dyes by actinomycetes. Microbiol. Lett., 107, 157-161.
- Zimmermann T., H.G. Kulla et T. Leisinger (1982). Properties of purified Orange II azoreductase, the enzyme initiating azo dyes degradation by Pseudomonas KF46. Eur. J. Biochem., 129, 179-203.
- Zissi U. et G. Lyberatos (1996). Azo-dye biodegradation under anoxic conditions. Water Sci. Technol., 34, 495-500.
- Zollinger H. (1987). Colour chemistry-synthesis, properties and applications of organic dyes and pigments. VCH Publishers Inc., New York, NY, USA.
Liste des figures
Figure 1
Exemples des groupes chromophores et auxochromes des colorants de types azoïques et anthraquinones.
Example of the chromophoric and auxochromic groups of azo dyes and anthraquinone.
Présentation d’un procédé d’ennoblissement textile (Hao et al. 2000).
Figure 2. Textile finishing process (Hao et al. 2000).
Figure 3
Mécanisme de dégradation de 4-(4’-sulfophénylazo)-2,6-diméthylephénole par la laccase de Pyricularia oryzae (Chivukula et Renganathan, 1995).
Figure 3. Degradation mechanism of 4-(4’-sulfophenylazo)-2,6-dimethylephenol by laccase from Pyricularia oryzae (Chivukula et Renganathan, 1995).
Figure 4
Réduction du rouge réactif 22 par Pseudomonas luteola (Chang et al., 2001).
Figure 4. Reduction of reactive red 22 by Pseudomonas luteola (Chang et al., 2001).
Figure 5
Minéralisation du colorant azoïque jaune mordant 3 (MY3) à l’aide d’un système de réacteur séquentiel anaérobie/aérobie (Haug et al., 1991).
Figure 5. Mineralization of azo dye mordant yellow 3 (MY3) by the sequential anaerobic/aerobic reactor (Haug et al., 1991).
Figure 6
Mécanisme de dégradation de l’acide orange 20 par Pseudomonas K24 (Kulla et al., 1983).
Figure 6. Degradation mechanism of acid orange 20 by Pseudomonas K24 (Kulla et al., 1983).
Liste des tableaux
Tableau 1
Principaux groupes chromophores et auxochromes classés par intensité croissante.
Table 1. Principle chromophoric and auxochromic groups classified by increasing intensity.
Tableau 2
Colorants azoïques révélés mutagènes et/ou carcinogènes.
Table 2. Azo dyes revealed mutagen and/or carcinogen.
Tableau 3
Taux de fixation sur la fibre textile pour les différentes classes tinctoriales de colorants azoïques (O’Neill et al., 1999; Azbar et al., 2004).
Table 3. Rate of fixation on the textile fiber for the various tinctoriales classes of azo dyes (O’Neill et al., 1999; Azbar et al., 2004).
Tableau 4
Comparaison de technologies physiques et chimiques, de dépollution des effluents textiles chargés de colorants synthétiques (Galindo, 1998).
Table 4. Comparative study of physical and chemical technology for the depollution of textile effluents charged by the synthetic dyes (Galindo, 1998).
Tableau 5
Quelques études sur la réduction de colorants azoïques par des bactéries aérobie incubées dans des conditions limitées en oxygène.
Table 5. Some studies on the azo dyes reduction by anaerobic bacteria incubated in the limited oxygen conditions.
Tableau 6
Principaux travaux portant sur la biodégradation des colorants azoïques dans des conditions oxygénées.
Table 6. Main works concerning the biodegradation of azo dyes in oxygenated conditions.