
Résumés
Abstract
Diatom analyses of sediments from a high elevation lake situated in an Engelmann Spruce - Subalpine Fir (ESSF) forest of south-central British Columbia, Canada, reveal long-term climate and water chemistry change. During the transition from the late-glacial / Pleistocene to the xerothermic early Holocene, small, benthic Fragilaria diatoms species that grew under low light conditions in Sicamous Creek Lake gave way to planktonic Cyclotella species that require open-water conditions. Warm temperatures in the mesothermic Holocene are indicated by smaller Cyclotella species and large, benthic pennate diatoms. Diatom communities reflected Neoglacial cooling in the late Holocene, with abundant Nitzschia fonticola and Achnanthes minutissima. Small, benthic Fragilaria regained abundance, suggesting cooling and conditions similar to the late-glacial interval. Diatom community composition responded to the deposition of the Mt. Mazama and Mt. St. Helens tephras, though the Mazama eruption caused greater change in relative abundance of various taxa within the assemblage. Correspondence analysis shows distinct communities have occurred since the initiation of sedimentation, likely due to climate controlled landscape and vegetation changes; diatom-inferred pH values using various models and training sets show limited acidification change occurred through the lake’s history.
Keywords:
- ESSF,
- diatom,
- transfer function,
- Holocene,
- climate change,
- correspondence analysis
Résumé
Les analyses de diatomées fossiles préservées dans les sédiments du lac Sicamous Creek en haute altitude, situé dans la partie centrale-sud de la Colombie-Britannique, Canada, révèlent des changements du climat et de la chimie de l'eau dans une forêt composée principalement d’épinette d’Engelmann et de sapin subalpin. Au passage du Tardiglaciaire / Pléistocène à l’Holocène supérieur xérothermique, les petites espèces benthiques du genre Fragilaria qui dominaient dans le lac Sicamous Creek sous des conditions de faible lumière ont été remplacées par des espèces planctoniques du genre Cyclotella qui sont favorisées par des conditions d'eau ouverte. Les températures élevées durant l’Holocène mésothermique sont caractérisées par des espèces de Cyclotella de petite taille et par des diatomées pennées benthiques de grande dimension. Le refroidissement dû à la néoglaciation durant l’Holocène inférieur est reflété par l’abondance de Nitzschia fonticola et Achnanthes minutissima dans les communautés de diatomées. Les petites diatomées benthiques du genre Fragilaria regagnaient en abondance, suggérant des conditions froides semblables à celles observées durant le Tardiglaciaire. La composition de la communauté des diatomées a changé en réponse aux retombées aériennes de téphra en provenance des monts Mazama et St. Helens, bien que l'éruption du mont Mazama ait provoqué un plus grand changement dans l’assemblage. L'analyse de correspondances démontre l’apparition des communautés distinctes depuis le début de la sédimentation probablement en raison des changements du paysage et de la végétation qui étaient contrôlés par le climat. Des valeurs de pH déduites des diatomées, utilisées dans différents modèles statistiques et avec diverses séries de calibration, indiquent une acidification limitée au cours de l'histoire du lac.
Mots-clés :
- Diatomées,
- fonction de transfert,
- Holocène,
- changements climatiques,
- l'analyse de correspondances
Corps de l’article
1. Introduction
Environmental conditions at high latitudes or at high elevations in mid-latitudes are harsh, as abundant snow and cold temperatures limit aquatic plant growth to a short ice-free period. In such settings, diatoms (class: Bacilliariophyceae) may be at their ecological limit (e.g. GOUDSMIT et al., 2000, PERREN et al., 2003, RÜHLAND and SMOL, 2005), and thus are particularly sensitive to changes in the physical and chemical parameters of the surrounding water (DOUGLAS and SMOL, 2010, LOTTER et al., 2010), which is in turn influenced by climate, though local climates may also be influenced by other factors, such as altitude, aspect, and topography (GULLET and SKINNER, 1992). Because of their good preservation, fast growth rates, and direct response to environmental changes, diatoms from subalpine lakes are powerful tools in paleolimnological analyses (MOSER, 2004) providing detailed records of past environmental change (BATTARBEE, 2000, HOBBS et al., 2010). Direct human influence is often negligible at remote subalpine lakes, thus paleoecological investigations are likely to yield reconstructions driven largely by climatic rather than other natural or anthropogenic processes (LOTTER et al., 2010), except where records are either very short or disturbed by natural events, such as landslides (MOSER, 2004).
Studies of aquatic changes at higher elevations in southern British Columbia have been rare (CHASE et al., 2008, GAVIN et al., 2011), relative to the number and diversity of lakes present (SCHIEFER and KLINKENBERG, 2004). Few of these studies have had environmental variables reconstructed using diatoms, as most diatom studies in British Columbia have examined anthropogenic influence upon lake ecosystems (CUMMING and LAIRD, 2006, DREGER, 2001, HEINRICHS et al., 2005, REAVIE et al., 2000) or long-term climate change from low-elevation sites (e.g. BENNETT et al., 2001, HEINRICHS and WALKER, 2006). The widely accepted general trend for postglacial climatic change in southern British Columbia begins with a cool late glacial period, followed by warm temperatures in the early- to mid-Holocene, and followed by a trend towards increased available moisture and decreasing temperatures in the late Holocene (CHASE et al., 2008, HEBDA, 1995, WALKER and PELLATT, 2003, 2008).
The relationship between diatom distribution and ecoclimatic or vegetation zones (sensu LOTTER et al., 2010) suggests that for British Columbia, different aquatic communities are expected to occur in alpine tundra compared to the subalpine zone and a montane forest, and thus vary through differing climatic episodes (GAVIN et al., 2011). With changes of climate, soil, or vegetation through time, concomitant changes in water pH and dissolved organic carbon (DOC) are also expected (BATTARBEE, 1990, PIENITZ and SMOL, 1993). RENBERG (2000) suggested that natural acidification from 12,000 to 2,300 BP occurred in lakes of southern Sweden due to soil acidification, decreasing base cations, and nutrient flux with developing vegetation. PRATHER and HICKMAN (2000) identified a similar natural acidification trend in a boreal forest lake from northeastern Alberta.
Confounding events of ecoclimatic diatom assemblages may be rare, but significantly large, volcanic tephra influx has affected diatom community relative abundance and composition in lakes elsewhere (BARKER et al., 2000, 2003, BARSDATE and DUGDALE, 1972, BERTRAND et al., 2005, BIRKS and LOTTER, 1994, CRUCES et al., 2006, KILIAN et al., 2006, LOTTER et al., 1995, STEPHENS et al., 2012, TELFORD et al., 1999, 2004, URRUTIA et al., 2007, VOLLAND 2006, WISSMAR et. al., 1982).
The present study at Sicamous Creek Lake focuses on the following objectives:
determine past aquatic environments based on qualitative changes in fossil diatom assemblages;
quantitatively examine the diatom record through ordination analysis and reconstruct the lake water chemistry using diatom transfer functions;
relate any trends or states with known patterns of regional Holocene climate change, and
examine the diatom record for a natural acidification trend and responses to volcanic tephra inputs.
2. Material and methods
2.1 Study site
The study site is located in the Engelmann Spruce - Subalpine Fir (ESSFwc (wet cold)) biogeoclimatic subzone (Figure 1) (MEIDINGER and POJAR, 1991), the uppermost forested zone in southern British Columbia. It occurs below the alpine tundra at elevations from 900 to 2,300 m. The ESSF exhibits a cold, moist and snowy continental climate with cool summers and long, cold winters. Mean annual temperatures vary between -2 to +2°C. Precipitation, which mostly falls as snow, varies highly in the zone; dry areas receive 400 to 500 mm, whereas wetter regions receive up to 2,200 mm.
Figure 1
British Columbia with the location of the training set lakes and the Shuswap region (inset). Magnification shows Sicamous Creek Lake (arrow) and the location of other nearby study sites in the ESSF.
La Colombie-Britannique avec la localisation des lacs de la série de calibration ainsi que la région de Shuswap (encadré). Le grossissement montre le lac Sicamous Creek (flèche) et la localisation d'autres sites d'étude à proximité dans l'ESSF.
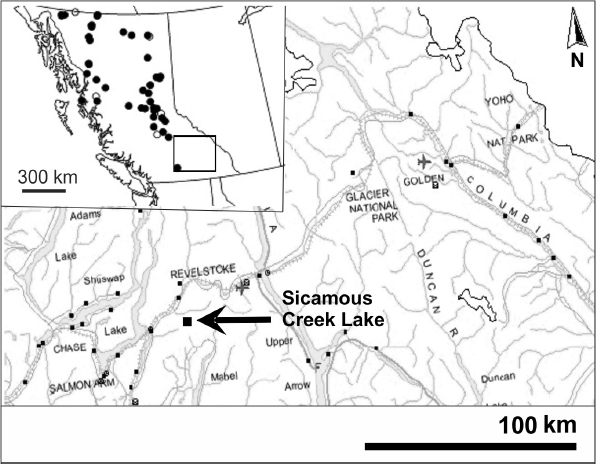
Sicamous Creek Lake (unofficially named) (50°, 50', 7" N; 118°, 48' 12" W; 1,690 m asl) is located approximately 10 km southeast of the town of Sicamous (Figure 1), in the Shuswap region of British Columbia. Climate data from Glacier National Park (1,323 m and 1,875 m), about 115 km from the site, recorded a mean annual temperature of 1.6°C and 0.2°C, respectively, and an annual precipitation over 1,500 mm and 2,000 mm, respectively (ENVIRONMENT CANADA, 2004). The watershed has approximately 50% cover of Subalpine fir (Abies lasiocarpa) (HEINRICHS et al., 2002). It is a shallow (Zmax = 1.9 m) small lake (2.4 ha), with a late summer pH value of 6.45, dissolved O2 value of 101.8% at 7.9°C, and conductivities ranging from 16.2 to 17.0 µS•cm-1 (sampled September 2005). Bedrock in the area is composed of granitic gneiss and the soils are sandy loam (PARISH et al., 1999). Abundant Carex, Scirpus, and Sphagnum are found along the surrounding lake margin, and yellow pond-lilies (Nuphar polysepela Engelm (Engelm.) E.O. Beal) extend several metres from shore towards the lake’s centre.
2.2 Field sampling
A 410 cm-long core was obtained from the deepest part of the lake using a modified Livingstone piston corer (WRIGHT, 1967) in the early autumn of 1998. The core was immediately sliced into 1, 2, 5 and 10 cm sections, depending upon stratigraphic markers and approximate periods of interest (e.g. early Holocene). Samples were taken for radiometric dating and the remainder of the core was stored until 2005, when it was processed for diatom analysis. A 20 cm-long core was also obtained in September 2005 using a Kajak-Brinkhurst corer to retrieve an undisturbed modern sediment sample.
2.3 Dating
The chronology in this study is based on two basal accelerator mass spectrometry (AMS) 14C dates from terrestrial macrofossils and two tephra layer dates (HEINRICHS et al., 2002); the radiocarbon dates from bulk sediment with a higher estimate error were omitted. The upper tephra layer is considered the Mt. St. Helens Yn tephra, dated to 3,390 ± 130 14C years BP (MULLINEAUX, 1996), based on its position below a bulk sediment date of 2,510 ± 40 14C years BP (HEINRICHS et al., 2002). The second tephra is volcanic ash from the Mt. Mazama eruption, considered to have occurred at 6730 ± 40 14C years BP (ZDANOWICZ et al., 1999). Calibrated years BP were calculated using CALIB 5.0 software (STUIVER and REIMER, 1993) extracting the 50% probability value from the cumulative probability graph.
2.4 Diatom analysis
2.4.1 Modern data
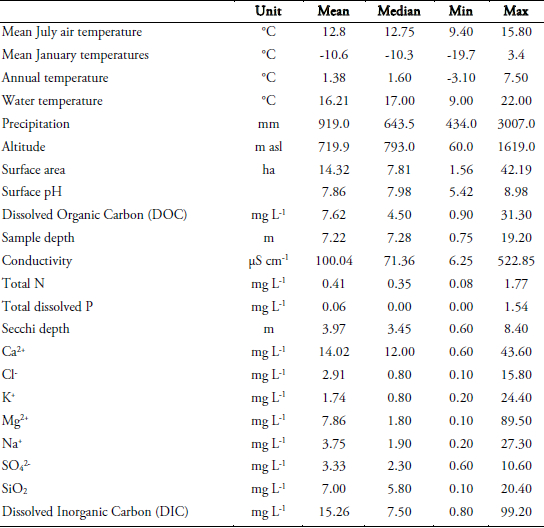
To assess the most significant environmental factors influencing diatom community composition, canonical correspondence analysis (CCA) using the program CANOCO version 3.12 (TER BRAAK, 1991) was performed on data (Table 1) derived from a 30 lake British Columbia diatom training set which includes 260 taxa (RACCA, unpublished, Figure 1). Only species with abundances greater than 1% and occurring in at least two samples were included. Species data were square-root transformed and rare species downweighted. The significance of the variables was tested using 999 unrestricted Monte Carlo permutations (TER BRAAK and ŠMILAUER, 1998). The relationship between species and the significant environmental parameters is illustrated in a detrended correspondence analysis (DCA) using CANOCO version 4.02 (TER BRAAK and ŠMILAUER, 1998) with square-root transformed species abundances and downweighting of rare species.
Table 1
Altitude, physical, chemical, climatic and morphometric variables of the 26 lakes in the British Columbian training set.
L’altitude et les variables physiques, chimiques, climatiques et morphométriques des 26 lacs faisant partie de la série de calibration des lacs de la Colombie-Britannique.
2.4.2 Fossil data
Diatoms were prepared after RENBERG (1990b) using approximately 0.01 g of dry or 0.1 g of wet sediment. Samples were treated with 30% H2O2 for four hours at 80°C until all organic material was digested. Excess base and carbonate, if present, were neutralised with 50% HCl, followed by three rinsing steps. A weak ammonia (NH3) solution was added to keep any clay in suspension and prevent diatom frustules from adhering to each other when making slides. Slides were prepared with 1-2 drops of a 1:10 or 1:20 dilution of the diatom suspension, dried on a cover slip, and mounted on slides using a 1:2 mixture of Naphrax and toluene at approximately 50°C.
Following BATTARBEE et al. (2001), a minimum of 400 diatom valves were counted at 10, 5, 2 or sometimes 1 cm intervals throughout the core. Broken valves that did not contain the centre, as well as girdle views were not included in the sum of valves. Identifications were made at a magnification of 1000x using an Olympus BX50 with reference mainly to KRAMMER and LANGE-BERTALOT (1991a,b, 1997a,b) and CUMMING et al. (1995). For identifications of taxa belonging to the genus Brachysira, LANGE-BERTALOT and MOSER (1994) was used.
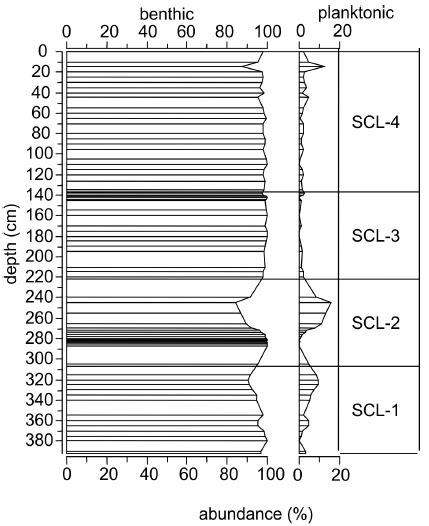
Taxon abundances were calculated as relative abundances of total identified diatoms. Only diatoms with abundances greater than 1% and occurrences in at least two samples were included in the stratigraphic diagram. Data (Figure 2) were compiled and graphed using C2 version 1.4 Beta (JUGGINS, 2003). Stratigraphically constrained sum-of-squares cluster analysis (CONISS) was applied to the square root transformed percent data of all species (GRIMM, 1987) to objectively divide the abundance data into zones. CONISS compares the samples of the different depths in terms of their similarity or dissimilarity and clusters similar data into zones. Relative abundances of planktonic vs. benthic taxa (Figure 3) were also compared to investigate possible shifts in lake level (WOLIN and DUTHIE, 1999). Species diversity was calculated with the Shannon-Weaver index, which measures the rarity or commonness of species in a community – the higher the number of taxa, the higher the diversity. Using the diversity index value, the taxa-independent evenness can be calculated using Pielou’s index, which indicates taxa abundance distributions by comparing maximal diversity to actual diversity. If all taxa are equally abundant, evenness is equal to one.
Figure 2
The diatom stratigraphic record for Sicamous Creek Lake. Taxa with a relative abundance greater than 1% and an occurrence in at least two samples are shown. Abundances are presented as a percentage of total identifiable diatoms.
La stratigraphie des diatomées fossiles du lac Sicamous Creek. Seuls les taxons avec une abondance supérieure à 1 % et qui sont présents dans au moins deux échantillons sont retenus. Les abondances représentent le pourcentage du total des diatomées identifiables.

Figure 3
Benthic versus planktonic diatoms from Sicamous Creek Lake. The uppermost interval (0 cm) is from 1998.
Les diatomées benthiques versus les diatomées planctoniques du lac Sicamous Creek. L'intervalle supérieur (0 cm) date de 1998.
A correspondence analysis (CA) using the program CANOCO version 3.12 (TER BRAAK, 1991) was performed on square-root transformed percentage abundances of fossil diatom species to show the correlation of species and samples, as well as the changes in diatom community through time. A total of 247 species from 26 lakes of the British Columbia diatom training set were used to perform the CA analysis.
To estimate past pH values, fossil diatom assemblages were compared against modern assemblages from 26 lakes of the British Columbia training set, with a pH range from 5 to 9.
Using a weighted averaging (WA) model in the program C2 version 1.4 Beta (JUGGINS, 2003), similarities were determined between fossil diatom communities of the Sicamous Creek Lake core and modern environmental and species data from the training set lakes. This WA model identified the more significant environmental factors selected from the CCA. WA assumes non-linear, unimodal species responses along environmental gradients. The WA-optimum of a taxon is the abundance-weighted mean of all its percentages in the record.
A chi-square test was performed on the uppermost interval of the Livingstone piston core and the uppermost five intervals from the Kajak-Brinkhurst sampler. Only taxa with non-zero values were compared.
3. Results and discussion
3.1 Environmental parameters
The lake showed nearly uniform temperatures and conductivity to maximum depth, indicating that light is able to penetrate to the lake bottom, and circulation had occurred. Dissolved oxygen values showed 100% saturation, confirming that the lake overturned in late summer, or is polymictic. Conductivity was low, possibly suggesting limited weathering of bedrock and soils and that little mineral matter enters the lake. At present, the lake water pH is circumneutral with a tendency towards acidity. This is typical for small, subalpine lakes (BAKER et al., 2012, CLOW et al., 2002, KOPÁČEK et al., 2000, LARSEN et al., 1999, NANUS et al., 2012, NYDICK et al., 2003). The abundant macrophytes provide extensive habitat for periphytic diatoms (VERMAIRE et al., 2011).
3.2 Diatom calibration - modern data
Canonical correspondence analysis (CCA) of the modern British Columbia diatom communities showed that the statistically most influential environmental variables (p<0.05) are pH (p=0.001), total dissolved phosphorus (P) (p=0.016), conductivity (p=0.017) and mean July air temperature (p=0.024). A total of 12 variables together with 110 species were analyzed. Variables having no measurably significant influence on diatom communities, like Na+ or Mg2+ concentrations, were excluded from the analysis.
Correlations were calculated between the detrended correspondence analysis (DCA) axes and the four significant environmental variables (surface pH, conductivity, mean July air temperature and total dissolved P) and 110 diatom taxa from 26 lakes (Figure 4, Table 1). The correlation matrix of the DCA (Tables 2 and 3) shows a high correlation between species and environmental axes 1 with pH (0.78 and 0.85) and conductivity (0.69 and 0.75). Conductivity and pH in turn are also positively correlated (0.69). Total dissolved P is negatively correlated with species and environmental axes 2 (0.43 and 0.73) as well as with environmental axis 4 (0.94), whereas it shows positive values in correlation with species and environmental axes 3 (0.49 and 0.84) and conductivity (0.4). Mean July air temperature is also highly negatively correlated with species and environmental axes 2 (0.46 and 0.79).
Figure 4
Detrended correspondence analysis (DCA) with four environmental variables, 26 lakes and 110 diatom species (abundance >1% and occurrence in at least two samples) from the British Columbian training set. Numbers represent diatom species (see Table 3), KW-numbers represent lakes, and environmental variable arrows are shown exaggerated (5x).
L'analyse ACD effectuée à partir de quatre variables environnementales, 26 lacs et 110 espèces de diatomées (abondance >1 % et apparition dans au moins deux échantillons) de la série de calibration des lacs de la Colombie-Britannique. Les numéros représentent les espèces de diatomées (voir le Tableau 3), les numéros KW représentent les lacs, et les flèches des variables environnementales ont été exagérées cinq fois.
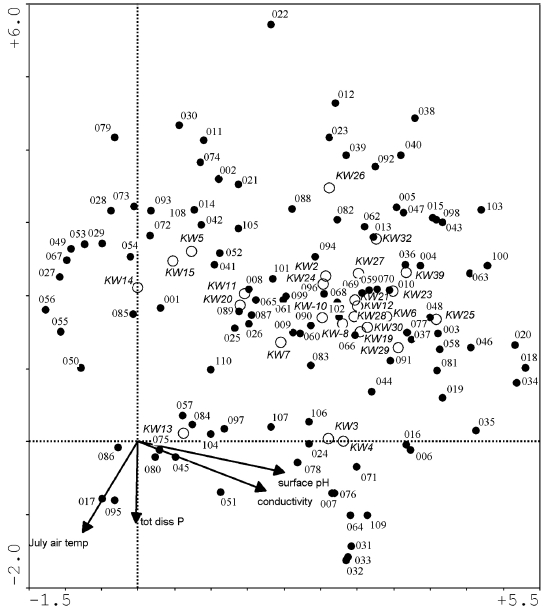
Tableau 2
Correlation matrix of environmental variables, resulting from detrended correspondence analysis (DCA) with the training set data.
La matrice de corrélation des variables environnementales, résultant de l’analyse des correspondances (ACD) avec les valeurs de la série de calibration.
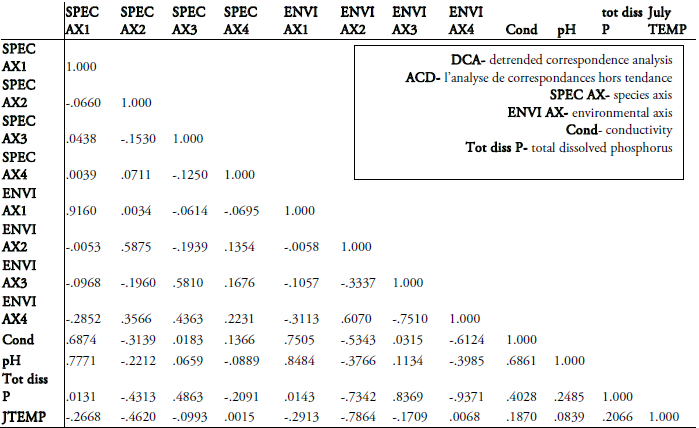
Tableau 3
Numbers for species occurring in detrended correspondence analysis (DCA) plot with training set data.
Les numéros des espèces figurant dans l’analyse ACD faite avec les valeurs de la série de calibration.

3.3 Chronology
The chronology of the sediment core calculated from radiocarbon and tephra dates spans a time frame from 1998, when the core was sampled, to 15,180 calibrated years BP. As grus was found at the core base, it is likely that the record spans the lake’s entire history since deglaciation. The first basal AMS date of 12,850 14C yr BP corresponds to the retreat of the Cordilleran Ice Sheet in south-central British Columbia (CLAGUE, 1991, RYDER et al., 1991). According to RYDER et al. (1991), it existed until 13,500 14C yr BP and extended above 2,300 m asl. Sicamous Creek Lake lies at an elevation of 1,690 m and would have been ice-free before lower elevation sites, though diatom growth may have been limited. Abundant diatom assemblages were first found in the sample directly above the one used for AMS dating; therefore the lake must have been free of permanent ice cover by 12,850 14C yr BP.
The unmixed tephra and coherent age to depth relationship of AMS and bulk sediment dates indicate relatively constant sedimentation, suggesting that the sediment was undisturbed over time. This is rather surprising, since the lake is in moose (Alces alces) habitat, and is shallow and small enough that it might have been frequented extensively (SHACKLETON, 1999, SPALDING, 1989). Tephra layers are helpful chronological markers in lake and soil stratigraphies, though caution in lake sediments must be employed, as downward movement of tephra in gyttja has been noted elsewhere (ANDERSON et al., 1984, BEIERLE and BOND, 2002).
3.4 Diatom stratigraphy - subfossil data
A total of 141 diatom taxa from 23 genera were identified in 71 samples from the sediment core. No diatoms were found in the lowermost five samples of the core (410-396 cm), with exception of one Cyclotella valve in the 400-398 cm interval (Figure 2). The number of total valves counted was on average higher in the 12 uppermost samples (minimum of 500 valves) than for the other intervals (minimum of 400 valves). CONISS identified four distinct diatom zones in the core sequence.
3.4.1 Zone SCL-1 (307-394 cm) (15,000-8,500 cal. yr BP) Fragilaria-Denticula-Navicula
Several taxa attain their highest abundance in this zone (e.g. Fragilaria elliptica, Denticula kuetzingii, Navicula cryptotenella). Denticula elegans, Nitzschia sinuata var. tabellaria, Cyclotella radiosa, Brachysira calcicola, Cymbella delicatula and Navicula aurora only appear in this first late-glacial interval. Fragilaria brevistriata and F. elliptica peak with an abundance of ca. 25%, though F. brevistriata nearly disappears at the level of 350 cm. F. elliptica has a second peak of about 40% after F. brevistriata disappears. F. elliptica declines to insignificant abundances towards the transition between zones SCL-1 and SCL-2. C. delicatula, D. kuetzingii and N. sinuata var. tabellaria rise to their maximum abundance throughout the core around 370-365 cm. At this level, N. aurora and Gomphonema angustatum increase – albeit to <10% abundance. Aulacoseira species are absent. The ratio of benthic versus planktonic diatoms in SCL-1 (Figure 3) is initially 97:3, passes through a 100:0 phase, decreases to 91:9, and finishes at a ratio of 92:8. Shannon diversity ranges from 1.6 to 2.4 (Figure 5b), evenness between 0.4 and 0.76 (Figure 5c) and richness varies from 23 to 45 (Figure 5d).
Figure 5
pH reconstruction with (a) British Columbian transfer function, (b) diversity, (c) evenness and (d) richness in Sicamous Creek Lake.
Les reconstitutions du pH avec (a) la fonction de transfert des lacs de la Colombie-Britannique, (b) la diversité, (c) l’uniformité et (d) la richesse du lac Sicamous Creek.

The diatom record from the beginning of this zone (394-360 cm), deposited during the late-glacial (>10,000 14C yr BP), is characteristic of cold conditions, as the small Fragilaria taxa are considered among the first diatoms to grow after marginal thawing of ice (LOTTER et al., 2010). Low diatom concentrations were also observed during counting of those intervals – although no absolute counts or concentrations were determined – indicating that lake productivity was either limited or that sedimentation rates were high. Cold temperatures, low nutrient concentrations, and high turbidity have been shown to inhibit diatom growth in north-east British Columbia during the late-glacial and early Holocene intervals (KARST-RIDDOCH et al., 2005).
The high variations in abundance of single taxa observed throughout this zone, such as the sudden decline (at 375-370 cm or 11,000 14C yr BP) and expansion (at 360-355 cm or ca. 9,800 14C yr BP) of F. elliptica and F. brevistriata, may have occurred in association with rapid and significant climatic changes correlated with the Younger Dryas (KOVANEN and EASTERBROOK, 2002, WALKER and PELLATT, 2003) or other significant climate events at ca. 9.3, 8.5, and 8.2 ka (GAVIN et al., 2011). Alpine glaciers expanded during the Younger Dryas (MENOUNOS and REASONER, 1997), which may have caused extended periods of ice cover resulting in the decline of planktonic diatom taxa (OHLENDORF et al., 2000). Diatom richness, evenness, and diversity fluctuations in the first 20 cm of the core support the idea of a rapidly changing environment.
The abundance of small, alkaliphilous Fragilaria taxa (F. elliptica, F. brevistriata) (SCHMIDT et al., 2004) suggests an elevated lake water pH during the late-glacial period. High diatom-inferred pH (DI-pH) supports this observation (Figure 5a). High pH values are expected with the input of groundwater or surface waters carrying elements of weathered soils and bedrock (SAULNIER-TALBOT et al., 2003, THOMPSON and DAUGHTRY, 2000) freshly exposed from glacial erosion.
The occurrence of planktonic Cyclotella (from 394-305 cm or 12,850-9,000 14C yr BP) suggests higher lake levels, likely due to increased snow / meltwater inputs, allowing more planktonic algae to grow (WOLIN and DUTHIE, 1999). An enhanced North American monsoon (BIRD and KIRBY, 2006) resulted in greater snowfalls likely earlier in winter, relative to today (ANDERSON et al., 2006). Given the winter solar radiation minima and summer maxima (GAVIN et al., 2011), rapid melting of this increased snow pack likely occurred. Cyclotella is described as a warm season dominant alga (STOERMER, 1977 as cited in HICKMAN and REASONER, 1994), and its presence may also support the interpretation of warmer summers, especially for the latter part of this zone.
The ratio of planktonic diatoms versus periphytic Fragilaria taxa in arctic and alpine lakes may reflect the length and extent of ice cover (DOUGLAS and SMOL, 2010, LOTTER and BIGLER, 2000, SMOL, 1988). As thawing starts from the lake edge, the littoral habitats become available for colonization by benthic flora first. Highly adaptable and considered as pioneers (HICKMAN and REASONER, 1994, PIENITZ et al., 1995), r-strategy flora consist mainly of small Fragilaria taxa, which are favoured under cold temperatures and low light conditions (LOTTER et al., 2010, LOTTER and BIGLER, 2000). Planktonic diatoms are only able to grow when the entire lake is ice-free. At Sicamous Creek Lake during the Pleistocene - Holocene boundary (10,000 14C yr BP), the diatom community is less dominated by benthic forms (e.g., Fragilaria taxa) and therefore the increasing trend of planktonic forms observed near the end of this zone (early Holocene) suggests a longer ice-free period. Around 9,000 cal. yr BP, summer solar insolation maxima and winter minima likely resulted in significant lake ice cover, except that warmer summer temperatures may have melted snow and ice faster, resulting in a shorter ice-cover period. Small Fragilaria taxa would be at a disadvantage under these conditions. HICKMAN and REASONER (1994) observed a similar pattern of early dominance and subsequent decline in small Fragilaria species between 7,000 and 3,000 cal. yr BP in Yoho National Park, British Columbia. The earlier disappearance of the small Fragilaria taxa at Sicamous Creek Lake may be due to an earlier retreat of the glaciers in this region (see early basal AMS date) compared to Yoho National Park.
Zone SCL-1 exhibits limnologically and likely climatically unstable conditions, as expected in the transition from the cold late-glacial to a warm early Holocene. The large range of correspondence analysis axis 1 sample scores, between 3.8 and 0.3 in the ecosystem trajectory (Figure 6), is evidence of this variability. Axis 1 explains 26% of the variation and its values are all positive in zone SCL-1. The ecosystem trajectory appears highly variable, but it occupies an exclusive space along the positive x-axis, though values appear to move from higher values at the core base to values approaching zero by the zone end. Interestingly, most of the species with the highest axis 1 scores in the species trajectory (Figure 7) only appear in zone SCL-1 (e.g. Nitzschia sinuata var. tabellaria 2.9, C. delicatula 2.0, B. calcicola 1.7, D. kuetzingii 1.6). N. sinuata var. tabellaria and N. aurora occur only in zone SCL-1, and have high axis 1 species scores in the CA.
Figure 6
Correspondence analysis (CA) ordination analysis of fossil diatom abundance showing the ecosystem trajectory, plotted on axes 1 and 2 of the fossil data. Axis 1 shows the sample scores from the first variable (λ = 0.26) and axis 2 gives the sample scores from the second variable (λ = 0.15).
L'ordination de l’analyse de correspondances (AC) de l’abondance des diatomées fossiles montre la trajectoire de l'écosystème, rapporté sur les axes 1 et 2 des données fossiles. L'axe 1 indique les résultats des échantillons pour la première variable (λ = 0.26) et l'axe 2 montre les résultats des échantillons pour la deuxième variable (λ = 0.15).
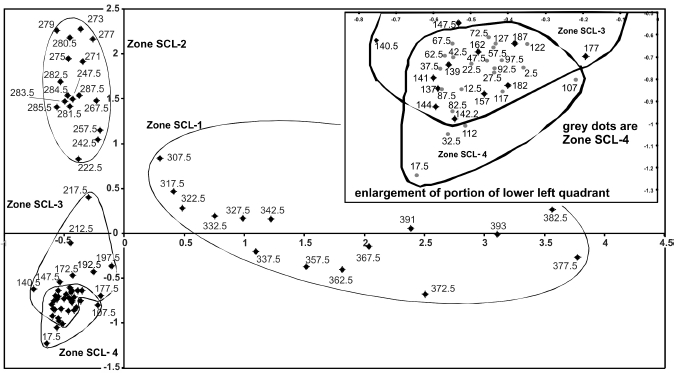
Figure 7
Correspondence analysis (CA): axis 1 versus axis 2 of the species scores.
L'analyse de correspondances (AC): l'axe 1 versus l'axe 2 des résultats des espèces.
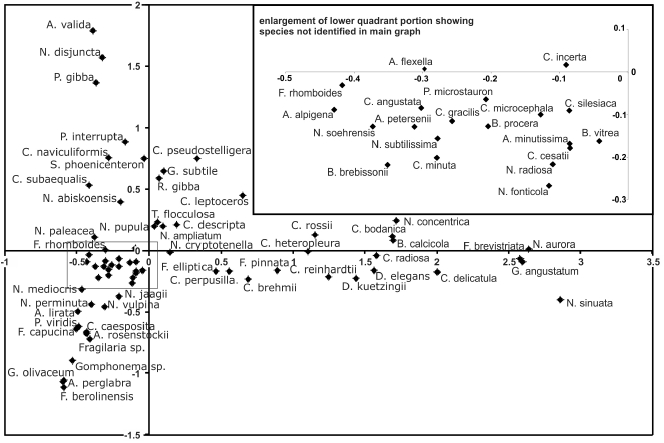
As pH was found to be amongst the three most significant environmental variables to explain diatom community changes in the CCA and appears correlated with species and environmental axes 1 in the DCA, pH is expected to have significant influence on axis 1 scores of the CA. Taxa with high axis 1 scores are likely alkalibiontic, and those with low axis 1 scores tolerant of more acidic conditions. Among the exclusively late-glacial species, only D. kuetzingii has a modern pH analogue in the British Columbia transfer function (pH optimum of 8.4; Figure 4). Here, the optima are located on the higher, more alkaline side of the pH scale and are considered as alkalibiontic species. ENACHE and PRAIRIE (2002) also found alkaline optima for C. delicatula and D. elegans in Québec surface sediment samples, supporting the alkaline optimum observed in the transfer function for these taxa. Since there are alkalibiontic taxa that occur in zone SCL-1 which do not appear in the British Columbian pH training set, even slightly higher diatom-inferred pH values can safely be assumed for zone SCL-1.
3.4.2 Zone SCL-2 (307-222 cm) (8,500-6,500 cal. yr BP) Pinnularia
This zone is characterized by Pinnularia interrupta, which occurs at 30 to 50% abundance. The larger Cyclotella taxa (e.g., C. radiosa) disappear in this level and give way to the smaller Cyclotella pseudostelligera that was also present in SCL-1, but shows approximately 15% abundance in SCL-2. A few Aulacoseira taxa, such as Aulacoseira valida, appear for the first time. Around 287-278 cm, the following taxa show peaks: Pinnularia gibba, Stauroneis phoenicenteron, and Tabellaria flocculosa. At the same time, the benthic community reaches a maximum abundance of 100%, as planktonic diatoms are completely absent from the lake. This distribution coincides with the deposition of Mt. Mazama ash. Nitzschia fonticola, C. pseudostelligera, and Achnanthes minutissima, as well as some Cymbella taxa that are under-represented in the tephra layer, increase, whereas P. gibba and S. phoenicenteron abundances drop immediately after the volcanic ash disturbance and stay low. P. interrupta and T. flocculosa continue to exhibit relatively high abundances. Navicula disjuncta appears only in this zone. After volcanic ash deposition, the planktonic diatom community increases again towards 3% abundance. Zone SCL-2 starts with low values for diversity and evenness, 1.54 and 0.40 respectively, but high values of richness at 47. After the Mazama ash deposition (278-280 cm), all three variables culminate, followed by an immediate decline (richness from 46 to 39). Diversity reaches its highest value at the level of 274-272 cm. Richness peaks 28 cm later (ca. 800 14C yr) with a maximum number of 53 taxa.
In contrast to the preceding zone, the composition and changes in diatom community suggest relatively warm and stable conditions which include the last 2,000 years of the xerothermic period and part of the transitional mid-Holocene (HEBDA, 2007). Large benthic pennate diatoms, such as Pinnularia sp., Stauroneis phoenicenteron, as well as Frustulia rhomboides usually occur with relatively warm temperatures in sub-arctic or subalpine regions (ROSÉN et al., 2000, SCHMIDT et al., 2004). Their abundances are higher in Zone SCL-2 compared to the previous zone, whereas smaller benthic pennates, like Fragilaria elliptica or N. fonticola, are less abundant. The dominance by benthic diatoms indicates relatively shallow water conditions (KARST-RIDDOCH et al., 2005, WOLIN and DUTHIE, 1999), reflecting the dry climate that prevailed at that time. Note that warming in large, deep lakes may favour planktonic species (sensu WINDER et al., 2009) due to changes in water clarity or thermal stratification, however Sicamous Creek Lake is more similar in size and climate to those examined by SCHMIDT et al. (2004) and ROSÉN et al. (2000).
The major event influencing diatom composition in this zone was deposition of the Mt. Mazama tephra at 280-278 cm, as biodiversity indicators changed dramatically. A local minimum in species richness after the tephra layer indicates that not all taxa were able to adapt to the post-depositional aquatic conditions. Another obvious consequence of the ash deposition is the higher abundance of P. interrupta during and especially immediately after the tephra layer. Many taxa of the genus Pinnularia are heavily silicified, suggesting that Si is a limiting nutrient for this genus, and their bloom here is likely due to the abundance of Si present in the ash (DRUITT and BACON, 1989). Frequent broken diatom valves were observed during counting, suggesting taphonomic processes may have affected the diatom assemblage, because breakage likely favoured the preservation of the more heavily silicified diatom valves and the smaller species. The significant change (from 1.7 to 2.2) in the CA axis 2 score versus depth plot at the time of the Mazama ash deposition (280-278 cm) supports the idea that ash deposition influenced the diatom community composition.
HICKMAN and REASONER (1994) speculate that tephra deposition in a lake may influence the benthic community by completely covering it, limiting light penetration and reducing the photosynthetic rate. However, the input of Si with the tephra could trigger higher productivity (SOMMER, 1994), affecting the Si:P ratio (HABERLE et al., 2000, ZIELINSKI, 2000), such that changes in diatom communities could be expected. These mechanisms are suspected to have occurred with the diatoms affected by Mazama ash in Lake Washington, Washington, USA (ABELLA, 1988) and Mahoney Lake, British Columbia, Canada (HEINRICHS et al., 1997).
Several studies document a climatic aftermath of large volcanic eruptions, and two to three years after the Mazama eruption, an average temperature reduction in the order of 0.6 to 0.7°C and/or a major decrease in stratospheric ozone are estimated to have occurred in the northern hemisphere (ZDANOWICZ et al.,1999). BRIFFA et al. (1998) note that over the past 600 years, certain localized cold summers coincide with large, single volcanic eruptions.
The presence of planktonic taxa such as C. pseudostelligera after deposition of the ash likely indicates continued warm water temperatures. Their occurrence at 270-220 cm (6,500-5,000 14C yr BP) falls within the moist and warm mesothermic period (7,000-3,500 14C yr BP) identified by HEBDA (1995, 2007). HICKMAN and REASONER (1994) found comparable abundances of one planktonic Cyclotella taxon at a similar time period (10,000 until 5,000 cal. yr BP) in subalpine Mary Lake, British Columbia, Canada. ROSENBERG et al. (2004) inferred cooler mean July air temperatures for the mesothermic period compared to the previous xerothermic period, thus Cyclotella might be affected by other variables than mean annual temperature. As the mid-Holocene in southern British Columbia started warm and ended with cooler temperatures (HEBDA, 2007), the decline of Cyclotella taxa at 5,000 14C yr BP potentially indicates the onset of cooling at this site.
The CA sample scores show axis 1 values change to negative values at the boundary with zone SCL-2 (Figure 6). Axis 2 sample score values explain 15% of the variation and are positive in this zone, likely due to the dominant presence of the influential P. interrupta, which has a species score of nearly 0.9.
The diatom-inferred pH estimates are slightly less alkaline than in zone SCL-1 and remain relatively stable. This pattern could be the beginning of a natural acidification trend (PENNINGTON, 1984, SAULNIER-TALBOT et al., 2003, SMOL, 2002) as regional ESSF vegetation with its relatively thick organic soil developed (HEINRICHS et al., 2002). Compared to zone SCL-1, changes in the diatom record in SCL-2 are mainly due to the environmental variable represented by axis 2 in the CA. The ecosystem trajectory plot shows that Zone SCL-2 has the highest positive values on axis 2.
3.4.3 Zone SCL-3 (222-137 cm) (6,500-3,550 cal. yr BP)
This zone is characterized by A. minutissima and N. fonticola, with abundant Achnanthes petersonii, Brachysira taxa, Cymbella incerta, Cymbella microcephala, Cymbella minuta, Cymbella silesiaca, and N. cryptotenella. All of these taxa were present in SCL-2, but in lower abundances. Diatoms of the genus Cyclotella disappear in this zone and Aulacoseira taxa continue with low abundances. P. interrupta, S. phoenicenteron and T. flocculosa decrease from 20 to <5% abundance after the first third of the zone. Only A. petersonii, Brachysira vitrea and N. fonticola exhibit higher abundances around the Mt. St. Helens tephra layer at 142.5-142 cm than shortly before. The benthic diatom community increases in proportion to the planktonic taxa in the previous zones, at approximately 98:2. The trend returns to fewer taxa, higher diversity and higher evenness values in zone SCL-3. Diversity values fluctuate around 1.8, evenness around 0.5, and richness around 38. Following the Mt. St. Helens Yn ash (142.5-142 cm), richness increases and values for diversity and evenness drop.
This zone falls into the second part of the mesothermic mid-Holocene and the beginning of the cooler late Holocene. In contrast to the preceding zone, the smaller pennate diatoms that dominate the assemblage, such as A. minutissima and N. fonticola, appear to be more competitive than the large benthic pennate diatoms, e.g. P. interrupta, Navicula pupula var. pupula and S. phoenicenteron, which may be in response to a decline in temperature (PIENITZ et al., 1995, ROSÉN et al., 2000, SCHMIDT et al., 2004). The decrease in P. interrupta at 215-210 cm (ca. 5,800 cal. yr BP) may also be due to lower temperatures or Si:P ratio, which can also influence diatom assemblages (KILHAM et al., 1986). CRAWFORD et al. (2001) determined that Pinnularia viridis is heavily silicified, and thus the Pinnularia genus may require high Si concentrations. Under normal circumstances, an early ice-out date and prolonged spring overturn leads to higher turbidity, which supplies heavily silicified diatoms with Si (KILHAM et al., 1996). The decrease in taxa of the genus Pinnularia in this zone may indicate a shorter ice-free period with a later ice-out date, consistent with mid-Holocene cooling. The zone coincided with the development of a dense macrophyte community, as the genus Brachysira is generally considered periphytic, and is associated with low or moderate conductivities (MCCORMICK, 2011). B. vitrea and Brachysira neoexilis require significant light intensity, typical of shallow water (GOOS, 2002), thus their presence here is not surprising, as Sicamous Creek Lake is now less than 2 m deep. The increase in N. fonticola, also periphytic, confirms the presence of dense macrophyte growth as a substrate.
Diatom-inferred pH changed little within this zone. The ecosystem trajectory plot shows Zone SCL-3 also occupies a unique space relative to the two preceding zones (Figure 6), suggesting the diatom community differed from those which occurred earlier. Note that fewer extreme values occur in axes 1 and 2 sample scores in this zone than in zones SCL-1 and SCL-2 (Figure 6), confirming the establishment of a stable aquatic environment. Thus it appears that less aquatic ecosystem disturbance is correlated with deposition of tephra from the Mt. St. Helens event than with the Mazama fall-out. As the CA change between SCL-2 and SCL-3 is a result of environmental variables influencing axis 2, the new equilibrium is likely not a result from a change in pH, but rather a combination of total P, conductivity, or temperature. These results suggest a stable limnological state or a new equilibrium. Low diversity, evenness, and moderate richness scores in SCL-3 compared to the previous zone are even more puzzling, as stable habitats should produce moderate richness with high evenness (DEATH, 1996). The new stable state is consistent with the apparent lack of change observed in the aquatic records of Cabin Lake and Lake of the Woods, British Columbia, Canada during this time (PALMER et al., 2002). This may be in part due to the lake being near the middle of an aquatic ecotone (sensu CHASE et al., 2008), though change is observed at this time in Yoho National Park, Canada (HICKMAN and REASONER, 1994). These results might also be interpreted as a disparity in regional climate trends or suggest a boundary to the aquatic ecotone.
Deposition of the Mt. St. Helens tephra at 142.5-142 cm (ca. 3,620 cal. yr BP) had less of an impact on the diatom community compared to the Mazama ash, probably due to the thinner layer deposited (0.5-1 cm versus 2-3 cm). The increases in A. petersonii, B. vitrea (tolerant to high light intensity) and N. fonticola after the Mt. St. Helens ash deposition likely reflect the decline in A. minutissima, which could have resulted from a loss of macrophytes. Species richness increases after the Mt. St. Helens tephra, unlike after the Mazama tephra, indicating that the two ash-depositing events did not have the same impact on the diatom community. It is interesting to consider whether the differences arose from the volume of tephra, timing of deposition, differences in chemistry, or perhaps even from secondary influences from an impact on terrestrial ecosystems in the catchment. Quantitative estimates of pH change vary, as the pH reconstruction indicates a decrease within two samples, from 7.9 to 7.7, after the St. Helens tephra.
Possible evidence of pH changes after ash deposition is observed in the CA ecosystem trajectory plot, as the sample situated immediately after the volcanic impact (140.5 cm) in zone SCL-3 is located furthest on the left side of axis 1, suggesting a decrease in pH in accordance with the British Columbian reconstruction. However, BATTARBEE et al. (2010) note that B. vitrea, which increases after the tephra deposition, is indicative of circumneutral conditions. No similar evidence for pH changes could be found in the CA for Mazama ash, which again highlights the difference in the two tephra events.
3.4.4 Zone SCL-4 (137-0 cm) (3,550 cal. yr BP - present)
This zone is characterized by 13 - 25% A. minutissima, 10% F. elliptica, and 20% N. fonticola. Brachysira brebissonii, B. procera, various Cymbella taxa, and N. cryptotenella are also present. The percentages of Aulacoseira taxa, Brachysira sp., Cymbella gracilis, Cymbella angustata, C. incerta, C. microcephala, C. minuta, C. silesiaca and N. cryptotenella are relatively stable. A. petersonii and B. vitrea both decrease early in the zone to trace amounts, and F. elliptica reappears reaching its maximum value of 32% late in the zone, falling to 11% at the zone end. Benthic diatoms dominate the zone, though at 15-10 cm, planktonic diatoms make up 13% of the total community. In zone SCL-4, diversity estimates fluctuate around 1.77 and evenness around 0.48. Richness varies between 37 and 50.
According to the CA ecosystem plot, zone SCL-4 shares many properties with zone SCL-3, although CONISS identified them as two different zones, likely because of differences in abundances of B. vitrea and F. elliptica. Despite their similarities, zone SCL-4 has some unique features compared to SCL-3. For example, the increased dominance of small Fragilaria taxa after 115-110 cm (ca. 3,000 cal. yr BP) coincides with the onset of cooler conditions of the late Holocene. These r-strategists are able to grow and reproduce when the lake thaws marginally only (LOTTER and BIGLER, 2000). As in the late glacial of zone SCL-1, small Fragilaria diatoms have an advantage over less cold-adapted benthic species. The significant, occasional decreases in the benthic diatom F. elliptica may reflect slightly warmer intervals during the prevailing cold conditions; however, not all benthic forms (e.g., N. fonticola) show change, suggesting that the conditions were generally cool. One peak of F. elliptica at 20-25 cm coincides with the maximum of the Little Ice Age in the 18th or 19th century (LUCKMAN, 2000). The decrease in B. vitrea, often associated with high light intensity and low nutrient ratios, suggests floating macrophytes (i.e. Nuphar) may have increased to the point where shading occurred. This interpretation is supported by periphyton increases, e.g. N. fonticola and the increase in the relative abundance of A. minutissima (VERMAIRE et al., 2011). Diatom-inferred pH shows a limited, gradual acidification trend in this zone, which would be expected with increasing organic matter accumulation (SAULNIER-TALBOT et al., 2003).
3.5 Modern uppermost sediment (2005 Kajak Brinkhurst core) (6-0 cm)
Characteristic taxa are A. minutissima with abundances between 7 and 12%, B. vitrea between 8 and 13%, N. cryptotenella up to 14% and N. fonticola with a maximum of 22% (BOROWSKI, 2005). Cymbella taxa and F. rhomboides display stable, but lower abundances. Small diatoms of the genus Fragilaria, like F. elliptica, and Aulacoseira taxa abundances are less than 1%. Benthic diatoms reach between 99 and 100% of the community. The uppermost sample from SCL-4 is not significantly different from the five top-most samples obtained using the Kajak Brinkhurst sampler (chi square test 1.00). This suggests that environmental impacts associated with recent forest harvesting and road building did not have an impact on the lake chemistry. This may be due to the extensive riparian area which was not disturbed, at least for several tens of metres from any harvesting activity, in concert with the limited gradient of the surrounding catchment.
3.6 Ordination analyses - fossil data
The plot of axes one and two of the species data from the CA (Figure 7) shows N. sinuata var. tabellaria at >2.8 of the ordinate axis, suggesting that it is at the end of an unconfirmed yet ecologically important variable. This species has been recorded from such diverse environments as shallow, concrete, artificial ponds in Turkey (SEN and SONMEZ, 2006), mountain streams in Spain (PÉREZ et al., 2009), and streams from eastern Québec, Canada (LAVOIE et al., 2009), where it was identified as a pollution-intolerant indicator. As pH and conductivity were significant drivers of the DCA axis 1, it is likely that pH, in part at least, influences the CA ordinate axis. However, since only 68% of taxa from the CA were in the DCA, this relationship is uncertain. F. brevistriata and G. angustatum are also significantly influential in the CA, and F. brevistriata occurs at >3.0 on the DCA axis 1, supporting the interpretation of pH as a factor in the CA. The abscissa possibly represents another ecological variable(s) with Fragilaria berolinensis (<-1.1) and A. valida (>1.7) on its extremities.
3.7 pH reconstructions
3.7.1 pH reconstruction from the British Columbia transfer function
Of the four models that were developed for pH reconstruction, weighted averaging (WA) with classical deshrinking was selected as it showed the best performance (Tables 2 and 4). The predictive ability is assessed with the correlation coefficient between the measured and diatom-inferred pH. The bootstrapped correlation coefficient (r2Boot) was 0.47 and the RMSEP (root mean squared error of prediction) was 0.54. This model yielded a range of reconstructed values from 7.7 to 8.2 (Figure 5a). The observed versus estimated pH plot for the 27 BC lakes (Figure 8a) shows few low values for pH (<7), but those present are at all times overestimated. High values (>7.3) were more common, and generally slightly underestimated. The severity of this bias is apparent from the residuals plot (Figure 8b).
Tableau 4
Performance statistics of the British Columbia pH transfer functions.
La performance statistique des modèles de fonction de transfert du pH de la Colombie-Britannique.

Figure 8
(a) Observed versus inferred WA pH values and (b) plot of residuals.
(a) Valeurs WA déduites versus valeurs WA observées du pH et (b) ordination des résidus.

The pH reconstruction using this model shows an increase at the beginning of zone SCL-1 (394-307 cm) from 8.0 to its highest value of pH 8.2. Thereafter, the pH decreases through SCL-2 until a value of 7.8 at 247.5 cm, where pH appears to stabilize. Zone SCL-3 shows an overall slight decrease from 8.0 at the beginning of the zone to 7.8 by the zone end. In zone SCL-4, pH values around 7.8 are inferred. Overall, the pH reconstruction shows a stable pH, with a slight acidification trend from 8.2 in the early postglacial to 7.7 at the present-day.
A reconstruction with the species data from the upper six centimetres extracted in 2005 did not result in different pH values than the ones already obtained with the species from the older uppermost samples.
3.7.2 Why is there no apparent long-term natural acidification trend in Sicamous Creek Lake?
Natural acidification in lakes occurs when drainage is impeded, often by paludification (SMOL, 2002), or in alpine/subalpine lakes due to acid deposition (NANUS et al., 2005). This lake acidification trend has been observed in areas with limited relief in northern Sweden (RENBERG, 1990a), Britain (BATTARBEE et al., 1985, PENNINGTON, 1984), and Canada (GORHAM et al., 1986). The relatively small decrease in pH over time early in the sediment record at Sicamous Creek Lake (from 8.0 to 7.7) and little subsequent additional acidification is unlike what SMOL (2002) described. Natural acidification in lakes is said to be mostly due to vegetation development and change. BATTARBEE et al. (2010) clarify that “… the rate and extent of acidification is also moderated by vegetation changes, and the impact of vegetation change on soils”. Transient disturbance in the catchment vegetation, such as fire or windthrow of trees, can lead to decreases in acidity (BATTARBEE et al., 2010), though increased acidity is due to longer-term processes in the composition of the catchment vegetation. PROBST et al. (1999) found that poorly buffered acid springs occurred more frequently in acid granitic areas covered by conifers. These conditions occur at Sicamous Creek Lake, as the granitic gneiss bedrock has poor buffering capacities (STAUFFER, 1990), the area is covered by a subalpine fir forest (PARISH, 1997), and there is long-term accumulation of organic/vegetation matter and Sphagnum surrounding the lake. Despite these conditions, little natural acidification of lake waters is observed.
Spruce-fir forests developed regionally around Sicamous Creek Lake by ca. 4,000 cal. yr BP (HEBDA and BROWN, in prep.) and subalpine fir was present since ca. 5,000 cal. yr BP (HEINRICHS et al., 2002). Obviously, there have not been major vegetation changes since this time, with the possible exception of limited natural fires and recent timber harvesting (PARISH et al., 1999).
Recent logging activities (ca. 1994) in the Sicamous Creek area (MITCHELL, 1997) resulted in the stand around the lake being partly harvested, such that base cations could leach easier into the lake. This may be the reason for the actual measured pH of 6.8. Species composition of the uppermost sample of the newly taken core (probably deposited after 1994) is not significantly different from the samples before, suggesting that the harvesting might not have had any detectable influence on the lake.
An alternative explanation for the small decrease in DI-pH could be inadequacies in the British Columbia training set, as it consists of few lakes, unevenly distributed along a pH gradient between 5 and 9. There are 13 lakes with pH >8 in the training set, 16 lakes with circumneutral pH, whereas only one lake has a pH <6. The species WA estimates are therefore biased, resulting in an edge effect at the acidic end of the gradient (HALL and SMOL, 1999). This is rather obvious in the residual plot (Figure 8b), with overestimated low pH values and underestimated high pH values. The result is that its predictive power is mostly restricted to the circumneutral region. However, the British Columbia transfer function presented here has a reasonable correlation coefficient (r2 (BC) = 0.90) and compares well with other pH models (e.g. ROSÉN et al., 2000: 0.61, WECKSTRÖM et al., 1997: 0.91).
4 Conclusion
The Sicamous Creek Lake diatom record, the first from the high-elevation Shuswap region of British Columbia, provides a ca. 15,000 year history of climate-driven aquatic environmental change. Diatom community zone SCL-1 (ca. 15,000-8,500 yr BP) corresponds to the cold late-glacial and the warm early Holocene periods. Fluctuations in diatom abundances of single taxa and correspondence analysis indicate a shift from cool glacial conditions towards the xerothermic early Holocene. Indicators of cool climate were small benthic Fragilaria taxa (mainly F. elliptica) whereas indicators of the continued warming were planktonic Cyclotella taxa. The ongoing warm phase of the early- and mid-Holocene is reflected by the diatom assemblages of Zone SCL-2 (ca. 8,500-6,500 yr BP). Indicator species for high temperatures and/or thermal stratification are the small planktonic centric C. pseudostelligera and the large benthic pennate P. interrupta. Stable conditions and low nutrient influx characterize Zone SCL-3 (ca. 6,500-3,500 yr BP), when a climax diatom community consisting of N. fonticola and A. minutissima developed. Zone SCL-4 shares similar scores with Zone SCL-3 with respect to an ecosystem trajectory plot, indicating that aquatic conditions have remained stable since ca. 6,500 yr BP. This zone also has a dominant F. elliptica population, suggesting ongoing cooling, consistent with cooling trends observed regionally (GAVIN et al., 2011).
Diatom community composition responded to deposition of the Mazama and Mt. St. Helens tephras, though quantitatively and qualitatively differently, perhaps due to less input from the Mt. St. Helens volcanic eruption. Extreme values in diatom diversity, richness, and evenness occurring immediately after deposition of Mazama tephra, coincident with a predominantly benthic community and silicified taxa, suggest that this input of volcanic materials had a markedly significant influence on the lake likely by affecting the Si:P ratio. Modern diatom communities had no detectable response with timber harvesting around the lake itself. Generally, this aquatic system appears resilient to moderate external influences such as climate changes or disturbance, as Holocene pH was stable and ecosystem trajectories varied greatest in the late-glacial period of lake and vegetation establishment. Natural acidification appears to be limited in the lake basin despite its location in the subalpine region on cation-depleted granitic bedrock. Further research investigating the links in the timing of vegetation changes and fire history with changes in aquatic conditions could improve our understanding of terrestrial- aquatic dynamics in this ecotone. Additionally, improved autecological knowledge for significant species identified in the ordination analyses, i.e., an expanded training set, would improve the interpretation of diatom records.
Parties annexes
Acknowledgement
The authors would like to thank Christina Bleskie, Marianne Chase, Lydia Stepanovic, Erin Barley and Peter Rotheisler for their warm welcome and inspiring creativity. We thank Kendrick Brown for his assistance in sediment coring. Dr. Arnulf Melzer is gratefully acknowledged for allowing completion of ACV’s diploma in British Columbia, Canada. A portion of this research was funded through an Okanagan University College Grant in Aid issued to MLH and by a grant of the National Science and Engineering Research Council of Canada awarded to RP and JR. The sediment core and radiocarbon dating were obtained through the support of a Forest Renewal British Columbia grant to RJH.
Bibliographie
- ABELLA S.E.B. (1988). The effect of the Mt. Mazama ashfall on the planktonic diatom community of Lake Washington. Limnol. Oceanogr., 33, 1376-1385.
- ANDERSON L., M.B. ABBOTT, B.P. FINNEY and S.J. BURNS. (2007). Late Holocene Moisture Balance Variability in the Southwest Yukon Territory, Canada. Quat. Sci. Rev., 26, 130-140.
- ANDERSON R.Y., E.B. NUHFER and W.E. DEAN (1984). Sinking of volcanic ash in uncompacted sediment in Williams Lake, Washington. Science, 225, 505-508.
- BAKER G.M., G. MORET, C.C. CAUDILL and N. TALLENT-HALSELL (2012). Year-round hydrologic monitoring of subalpine lakes in Great Basin National Park. In: Rethinking Protected Areas in a Changing World: Proceedings of the 2011 George Wright Society Biennial Conference on Parks, Protected Areas, and Cultural Sites. Weber S. (Editor), Hancock, Michigan: The George Wright Society, pp. 32-36.
- BARKER P., R. TELFORD, O. MERDACI, D. WILLIAMSON, M. TAIEB, A. VINCENS and E. GILBERT (2000). The sensitivity of a Tanzanian crater lake to catastrophic tephra input and four millennia of climate change. Holocene, 10, 303-310.
- BARKER P., D. WILLIAMSON, F. GASSE and E. GIBERT (2003). Climatic and volcanic forcing revealed in a 50,000-year diatom record from Lake Massoko, Tanzania. Quat. Res., 60, 368-376.
- BARSDATE R.J. and R.C. DUGDALE (1972). Effects of volcanic ashfalls on chemical and sediment characteristics of two Alaskan lakes. J. Fish. Res. B. Can., 29, 231-236.
- BATTARBEE R.W, D.F. CHARLES, C. BIGLER, B.F. CUMMING and I. RENBERG (2010). Diatoms as indicators of surface water acidity. In: The Diatoms: Applications for the Environmental and Earth Sciences, 2ndEdition. STOERMER, E.F. and J.P. SMOL (Editors), Cambridge University Press., pp. 98-121.
- BATTARBEE R.W., R.J. FLOWER, A.C. STEVENSON and B. RIPPEY (1985). Lake acidification in Galloway: a palaeoecological test of competing hypotheses. Nature, 314, 350-352.
- BATTARBEE R.W. (2000). Palaeolimnological approaches to climate change, with special regard to the biological record. Quat. Sci. Rev., 19, 107-124.
- BATTARBEE R.W. (1990). The causes of lake acidification, with special reference to the role of acid deposition. Phil. Trans. R. Soc. Lond. B, 327, 339-347.
- BATTARBEE R.W., V.J. JONES, R.J. FLOWER, N.G. CAMERON, H. BENNION, L. CARVALHO and S. JUGGINS (2001). Diatoms. In: Tracking Environmental Change Using Lake Sediments, Volume 3: Terrestrial, Algal and Siliceous Indicators. SMOL J.P., H.J.B. BIRKS and W.M. LAST (Editors), Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 155-202.
- BEIERLE B. and J. BOND (2002). Density-induced settling of tephra through organic lake sediments. J. Paleolim., 28, 433-440.
- BENNETT J.R., B.F. CUMMING, P.R. LEAVITT, M. CHIU, J.P. SMOL and J. SZEICZ (2001). Diatom, pollen, and chemical evidence of postglacial climatic change at Big Lake, south-central British Columbia, Canada. Quat. Res., 55, 332-343.
- BERTRAND S., X. BOËS, J. CASTIAUX, F.CHARLET, R. URRITIA, C. ESPINOZA, G. LEPOINT, B. CHARLIER and N. FAGEL (2005). Temporal evolution of sediment supply in Lago Puyehue (Southern Chile) during the last 600 years and its climatic significance. Quat. Res., 64, 163-175.
- BIRD B.W. and M.E. KIRBY (2006). An alpine lacustrine record of early Holocene North American Monsoon dynamics from Dry Lake, southern California (USA) J. Paleolim., 35, 179-192.
- BIRKS H.J.B. and A.F. LOTTER (1994). The impact of the Laacher See Volcano (11 000 yr BP) on terrestrial vegetation and diatoms. J. Paleolim., 11, 313-322.
- BOROWSKI A. (2005). A diatom-based paleoenvironmental record of a subalpine lake in south-central British Columbia, Canada. Diplom thesis, Technische Universität München, Germany, 88 p.
- BRIFFA K.R., P.D. JONES, F.H. SCHWEINGRUBER and T.J. OSBORN (1998). Influence of volcanic eruptions on Northern Hemisphere summer temperature over the past 600 years. Nature, 393, 450-455.
- CHASE M., C. BLESKIE, I.R. WALKER, D.G. GAVIN and F.S. HU (2008). Midge-inferred Holocene summer temperatures in Southern British Columbia, Canada. Palaeogeogr., Palaeoclimatol., Palaeoecol., 257, 244-259.
- CLAGUE J.J. (1991). Quaternary glaciation and sedimentation. In: Geology of the Cordilleran Orogen in Canada. GABRIELSE H. and C.J. YORATH (Editors), Geological Survey of Canada, Geology of Canada, Vol. 4., Chap. 12, pp. 419-434.
- CLOW D.W., R.G. STRIEGL, L. NANUS, M.A. MAST, D.H. CAMPBELL and D.P. KRABBENHOFT (2002). Chemistry of selected high-elevation lakes in seven national parks in the western United States. Water, Air, Soil Pollut. Focus, 2, 139-164.
- CRAWFORD S.A., M.J. HIGGINS, P. MULVANEY and R. WETHERBEE (2001). Nanostructure of the diatom frustule as revealed by atomic force and scanning electron microscopy. J. Phycol., 37, 543-554.
- CRUCES F., R. URRUTIA, O. PARRA, A. ARANEDA, H. TREUTLER, S. BERTRAND, N. FAGEL, L. TORRES, R. BARRA and L. CHIRINOS (2006). Changes in diatom assemblages in an Andean lake in response to a recent volcanic event. Arch. Hydrobiol., 165, 23-35.
- CUMMING B.F. and K. LAIRD (2006). Assessment of changes in total phosphorus in Nukko Lake, B.C.: a paleolimnological assessment. Prepared for James Jacklin, B.C. Ministry of Water, Land and Air Protection., pp. 1-8.
- CUMMING B.F., S.E. WILSON, R.I. HALL and J.P. SMOL (1995). Diatoms from British Columbia (Canada) and their relationship to salinity, nutrients and other limnological variables. Bibl. Diatomol., 31, 1-207.
- DEATH R.G. (1996). The effect of habitat stability on benthic invertebrate communities: the utility of species abundance distributions. Hydrobiol., 317, 97-107.
- DOUGLAS M.S.V. and J.P. SMOL (2010). Freshwater diatoms as indicators of environmental change in the High Arctic. In: The diatoms: Applications for the Environmental and Earth Sciences, 2nd Edition. STOERMER, E.F. and J.P. SMOL (Editors), Cambridge University Press, pp. 249-266.
- DREGER L. (2001). A diatom palaeoecological assessment of human impacts on Shannon Lake, Westbank, British Columbia, Canada. Directed Studies Thesis, Okanagan University College, Kelowna, BC, Canada, 26 p.
- DRUITT T.H. and C.R. BACON (1989). Petrology of the zoned calcalkaline magma chamber of Mount Mazama, Crater Lake, Oregon. Contr. Mineral. Petrol., 101, 245-259.
- ENACHE M. and Y.T. PRAIRIE (2002). WA-PLS diatom-based pH, TP and DOC inference models from 42 lakes in the Abitibi clay belt area (Quebec, Canada). J. Paleolim., 27, 151-171.
- ENVIRONMENT CANADA (2004). Canadian Climate Normals 1971-2000. http://climate.weatheroffice.ec.gc.ca [consultation August 14, 2006].
- GAVIN D., A.C.G. HENDERSON, K.S. WESTOVER, S.C. FRITZ, M.J. LENG and F.S. HU (2011). Abrupt Holocene climate change and potential response to solar forcing in western Canada. Quat. Sci. Rev., 30, 1243-1255.
- GOOS C. (2002). Einfluß des Periphytons auf das Lichtklima submerser Makrophyten. Dissertation, Technische Universität München, Germany, 205 p.
- GORHAM E., J.K. UNDERWOOD, F.B. MARTIN, J.G. OGDEN III (1986). Natural and anthropogenic causes of lake acidification in Nova Scotia. Nature, 324, 451-453.
- GOUDSMIT G.-H., G. LEMCKE, D.M. LIVINGSTONE, A.F. LOTTER, B. MÜLLER and M.STURM (2000). Hagelseewli: a fascinating mountain lake – suitable for palaeoclimate studies? Verh. Int. Ver. Limnol., 27, 1013-1022.
- GRIMM E.C. (1987). CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci., 13, 13-35.
- GULLET D.W. and W.R. SKINNER (1992). The State of Environment Report – The State of Canada’s Climate: Temperature Change in Canada 1895-1991. Environment Canada, Minister of Supply and Services Canada, Catalogue No. En1-11/92-2E. pp. 11-17.
- HABERLE S.G., J.M. SZEICZ and K.D. BENNETT (2000). Late Holocene vegetation dynamics and lake geochemistry at Laguna Miranda, XI Region, Chile. Rev. Chilena Hist. Nat., 73, 655-669.
- HALL R.I. and J.P. SMOL (1999). Diatoms as indicators of lake eutrophication. In: The diatoms: Applications for the Environmental and Earth Sciences. Stoermer E.F. and J.P. Smol (Editors), Cambridge University Press. pp. 128-168.
- HEBDA R.J. (1995). British Columbia vegetation and climate history with focus on 6 ka BP. Géogr. Phys. Quatern., 49, 55-79.
- HEBDA R.J. (2007). Biodiversity: Geological history in British Columbia. Report on the status of biodiversity in British Columbia. The Biodiversity BC Technical Subcommittee, BC, Canada, 11 p.
- HEINRICHS M.L., S.E. WILSON, I.R. WALKER, J.P. SMOL, R.W. MATHEWES and K.J. HALL (1997). Midge- and diatom-based paleosalinity reconstructions for Mahoney Lake, Okanagan Valley, British Columbia, Can. Int. J. Salt Lake Res., 6, 249-267.
- HEINRICHS M.L., J.A. ANTOS, R.J. HEBDA and B.A. GREG (2002). Abies lasiocarpa (Hook.) Nutt. in the late-glacial and early-Holocene vegetation of BC, Canada, and adjacent regions in Washington. USA. Rev. Palaeobot. Palynol., 120, 107-122
- HEINRICHS M.L., B.F. CUMMING, K.R. LAIRD and J.S. HART (2005). Diatom-and chironomid- inferred eutrophication of Bouchie Lake, British Columbia. Water Qual. Res. J. Can., 40, 418-430.
- HEINRICHS M.L. and I.R. WALKER (2006). Fossil midges and palaeosalinity: potential as indicators of hydrological balance and sea-level change. Quat. Sci. Rev., 25, 1948-1965.
- HICKMAN M. and M.A. REASONER (1994). Diatom responses to late Quaternary vegetation and climate change, and to deposition of two tephras in an alpine and a subalpine lake in Yoho National Park, British Columbia. J. Paleolim., 11, 173-188.
- HOBBS W.O, R.J. TELFORT, H.J.B. BIRKS, J.E. SAROS, R.R.O. HAZEWINKEL, B.B. PERREN, E. SAULNIER-TALBOT and A.P. WOLFE (2010). Quantifying recent ecological changes in remote lakes of North America and Greenland using sediment diatom assemblages. PLoS ONE, 5, doi: 10.1371.
- Juggins S. (2003). C2 version 1.4. User Guide. Software for ecological and palaeoecological data analyisis and visualisation. University of Newcastle, UK. 69 p.
- KARST-RIDDOCH T.L., M.F.J. PISARIC D.K. YOUNGBLUT and J.P. SMOL (2005). Postglacial record of diatom assemblage changes related to climate in an alpine lake in the northern Rocky Mountains, Canada. Can. J. Bot., 83, 968-982.
- KILHAM P., S.S. KILHAM and R.E. HECKY (1986). Hypothesized resource relationships among African planktonic diatoms. Limnol. Oceanogr., 31, 1169-1181.
- KILHAM S.S., E.C. THERIOT and S.C. FRITZ (1996). Linking planktonic diatoms and climate change in the large lakes of the Yellowstone ecosystem using resource theory. Limnol. Oceanogr., 41, 1052-1062.
- KILIAN R., H. BIESTER, J. BEHRMANN, O. BAEZA, M. FESQ-MARTIN, M. HOHNER, D. SCHIMPF, A. FRIEDMAN and A. MANGINI (2006). Millennium-scale volcanic impact on a superhumid and pristine ecosystem. Geology, 34, 609-612.
- KOPÁÈEK J., E.STUCHLIK, V. STRAŠKRABOVÁ and P. PŠENÁKOVÁ (2000). Factors governing nutrient status of mountain lakes in the Tatra Mountains. Freshwat. Biol., 43, 369-383.
- KOVANEN D.J. and D.J. EASTERBROOK (2002). Extent and timing of Allerød and Younger Dryas age (ca. 12.5-10.0 14C kyr BP) oscillations of the Cordilleran Ice Sheet in the Fraser Lowland, Western North America. Quat. Res., 57, 208-224.
- KRAMMER K. and H. LANGE-BERTALOT (1997a). Süßwasserflora von Mitteleuropa. Bacillariophyceae. H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauer (Editors), Band 2/1, Gustav Fischer Verlag Jena, 876 p.
- KRAMMER K. and H. LANGE-BERTALOT (1997b). Süßwasserflora von Mitteleuropa. Bacillariophyceae. H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauer (Editors), Band 2/2, Gustav Fischer Verlag Jena, 610 p.
- KRAMMER K. and H. LANGE-BERTALOT (1991a). Süßwasserflora von Mitteleuropa. Bacillariophyceae. H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauer (Editors), Band 2/3, Gustav Fischer Verlag Stuttgart, 576 p.
- KRAMMER K. and H. LANGE-BERTALOT (1991b). Süßwasserflora von Mitteleuropa. Bacillariophyceae. H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauer (Editors), Band 2/4, Gustav Fischer Verlag Stuttgart, 437 p.
- LARSEN G., G. LOMNICKY, R. HOFFMAN, W.J. LISS and E. DEIMLING (1999). Integrating physical and chemical characteristics of lakes into the glacially influenced landscape of the northern Cascade Mountains, Washington State, USA. Environ. Manage., 24, 219-228.
- LANGE-BERTALOT H. and G. MOSER (1994). Brachysira – Monographie der Gattung. Bibliotheca Diatomologica, Band 29, J. Cramer in der Gebrüder Borntraeger Verlagsbuchhandlung, 212 p.
- LAVOIE I., P.B. HAMILTON, Y.-K. WANG, P.J. DILLON and S. CAMPEAU (2009). A comparison of stream bioassessment in Québec (Canada) using six European and North American diatom-based indices. Nova Hedwigia. Beiheft., 135, 37-56.
- LOTTER A.F., H.J.B. BIRKS and B. ZOLITSCHKA (1995). Late-glacial pollen and diatom changes in response to two different environmental perturbations: volcanic eruption and Younger Dryas cooling. J. Paleolim., 14, 23-47.
- LOTTER A.F., R. PIENITZ and R. SCHMIDT (2010). Diatoms as indicators of environmental change in sub-arctic and alpine regions. In: The Diatoms: Applications for the Environmental and Earth Sciences, 2nd Edition. STOERMER, E.F. and J.P. SMOL (Editors), Cambridge University Press, pp. 231-248.
- LOTTER, A.F. and C. BIGLER (2000). Do diatoms in the Swiss Alps reflect the length of ice cover? Aquat. Sci., 62, 125-141.
- LUCKMAN B.H. (2000). The Little Ice age in the Canadian Rockies. Geomorphol., 32, 357-384.
- McCORMICK, P.V. (2011). Soil and periphyton indicators of anthropogenic water-quality changes in a rainfall-driven wetland. Wetland Ecol. Manage., 19, 19-34.
- MEIDINGER D. and J. POJAR (1991). Ecosystems of British Columbia. Special Report Series No. 6, British Columbia Ministry of Forests, Research Branch, Victoria BC, 330 p.
- MENOUNOS B. and M.A. REASONER (1997). Evidence for Cirque Glaciation in the Colorado Front Range during the Younger Dryas Chronozone. Quat. Res., 48, 38-47.
- MITCHELL J.L. (1997). Harvesting costs and logistics at Sicamous Creek. Abstract: Sicamous Creek silvicultural system project: workshop proceedings. 24/1997, Ministry of Forest research program, British Columbia, p. 15.
- MOSER K.A. (2004). Paleolimnology and the frontiers of biogeography. Phys. Geogr., 25, 453-480.
- MULLINEAUX D.R. (1996). Pre-1980 tephra-fall deposits from Mount St. Helens, Washington. U.S. Geological Survey Professional Paper, 1563, 1-99.
- NANUS L., D.H. CAMPBELL and M.W. WILLIAMS (2005). Sensitivity of alpine and subalpine lakes to acidification from atmospheric deposition in Grand Teton National Park and Yellowstone National Park, Wyoming. U.S. Geological Survey Scientific Investigations Report, 2005–5023, 37 p.
- NANUS L., D.W. CLOW, J.E. SAROS, V.C. STEPHENS and D.H. CAMPBELL (2012). Mapping critical loads of nitrogen deposition for aquatic ecosystems in the Rocky Mountains, USA . Environ. Pollut., 166, 125-135.
- NYDICK K.R., B.M. LAFRANCOIS, J.S. BARON and B.M. JOHNSON (2003). Lake-specific responses to elevated atmospheric nitrogen deposition in the Colorado Rocky Mountains, U.S.A. Hydrobiologia, 510, 103-114.
- OHLENDORF C., C. BIGLER, G.-H.GOUDSMIT, G. LEMCKE, D.M. LIVINGSTONE, A.F. LOTTER, B. MÜLLERS and M. STURM (2000). Causes and effects of long periods of ice cover on a remote high Alpine lake. J. Limnol., 59 (Suppl. 1), 65-80.
- PALMER S.L., I.R. WALKER, M.L. HEINRICHS, R.J. HEBDA and G.G.E. SCUDDER (2002). Postglacial midge community change and Holocene palaeotemperature reconstructions near treeline, southern British Columbia (Canada). J. Paleolim., 28, 469-490.
- PARISH R. (1997). Age and size structure of the forest at Sicamous Creek. In: Sicamous Creek Silvicultural Project: Working Paper. HALLSTEDT C. and A. VYSE (Editors), Kamloops, British Columbia, Canada. pp. 16-31.
- PARISH R., J.A. ANTOS and M.-J. FORTIN (1999). Stand development in an old- growth subalpine forest in southern interior British Columbia. Can. J. For. Res., 29, 1347-1356.
- PENNINGTON W. (1984). Long-term natural acidification of upland sites in Cumbria: Evidence from post-glacial lake sediments. Fifty-second annual report for the year ended 31st March 1984, Ambleside, UK, Freshwater Biological Association, pp. 28-46.
- PÉREZ M.C., N.I. MAIDANA and A. COMAS (2009). Phytoplankton composition of the Ebro River estuary, Spain. Acta Botanica Croatica, 68, 11-27.
- PERREN, B.B., R.S. BRADLEY and P. FRANCUS (2003). Rapid lacustrine response to recent high Arctic warming: a diatom record from Sawtooth Lake, Ellesmere Island, Nunavut. Arct. Antarct. Alp. Res., 35, 271-278.
- PIENITZ R. and J.P. SMOL (1993). Diatom assemblages and their relationship to environmental variables in lakes from the boreal forest-tundra ecotone near Yellowknife, Northwest Territories, Canada. Hydrobiologia, 269/270, 391-404.
- PIENITZ R., J.P. SMOL and H.J.B. BIRKS (1995). Assessment of freshwater diatoms as quantitative indicators of past climate change in the Yukon and Northwest Territories, Can. J. Paleolim., 13, 21-49.
- PRATHER C. and M. HICKMAN (2000). History of a presently slightly acidic lake in northeastern Alberta, Canada, as determined through analysis of the diatom record. J. Paleolim., 24, 183-198.
- PROBST A., J.P. PARTY, C. FEVRIER, E. DAMBRINE, A.L. THOMAS and J.M. STUSSI (1999). Evidence of springwater acidification in the Vosges Mountains (north-east of France): influence of bedrock buffering capacity. Water Air Soil Pollut., 114, 395-411.
- REAVIE E.D., J.P. SMOL, I.D. SHARPE, L.A. WESTENHOFER and A.-M. ROBERTS (2000). Paleolimnological analyses of cultural eutrophication patterns in British Columbia lakes. Can. J. of Bot., 78, 873-888.
- RENBERG I. (1990a). A 12 600 year perspective of the acidification of Lilla Öresjön, southwest Sweden. Phil. Trans. Royal Soc. Lond., Ser. B, 327, 357-361.
- RENBERG I. (1990b). A procedure for preparing large sets of diatom slides from sediment cores. J. Paleolim., 4, 87-90.
- RENBERG I. (2000). Lake sediments as environmental archives. In: Environmental Changes in Fennoscandia During the Late Quaternary. SANDGREN P. (Editor), LUNDQUA Report 37, Department of Quaternary Geology, Lund University, Sweden. pp. 90-95.
- ROSÉN P., R. HALL, T. KORSMAN and I. RENBERG (2000). Diatom transfer functions for quantifying past air temperature, pH and total organic carbon concentration from lakes in northern Sweden. J. Paleolim., 24, 109-123.
- ROSENBERG S.M., I.R. WALKER, R.W. MATHEWES and D.J. HALLETT (2004). Midge-inferred Holocene climate history of two subalpine lakes in southern British Columbia, Canada. Holocene, 14, 258-271.
- RÜHLAND K. and J.P. SMOL (2005). Diatom shifts as evidence for recent sub-arctic warming in a remote tundra lake, NWT, Canada. Paleogeogr. Paleoclimatol. Palaeoecol., 226, 1-16.
- RYDER J.M., R.J. FULTON and J.J. CLAGUE (1991). The Cordilleran Ice Sheet and the glacial geomorphology of southern and central British Columbia. Geogr. Phys. Quat., 45, 365-377.
- SAULNIER-TALBOT, É., R. PIENITZ and W.F. VINCENT (2003). Holocene lake succession and palaeo-optics of a sub-arctic lake, northern Québec, Canada. Holocene, 13, 517-526.
- SCHIEFER E. and B. KLINKENBERG (2004). The distribution and morphometry of lakes and reservoirs in British Columbia: a provincial inventory. Can. Geogr., 48, 345-355.
- SCHMIDT R., C. KAMENIK, C. KAIBLINGER and M. HETZEL (2004). Tracking Holocene environmental changes in an alpine lake sediment core: application of regional diatom calibration, geochemistry, and pollen. J. Paleolim., 32, 177-196.
- SEN B. and F. SONMEZ (2006). A study on the algae in fish ponds and their seasonal variations. Int. J. Sci. Tech., 1, 25-33.
- SHACKLETON D.M. (1999). Hoofed Mammals of British Columbia, Volume 3 The Mammals of British Columbia. Royal British Columbia Handbook, UBC Press Vancouver, 268 p.
- SMOL J.P. (1988). Paleoclimate proxy from freshwater arctic diatoms. Ver. Int. Ver. Limnol., 23, 837-844.
- SMOL J.P. (2002). Acidification: finding the ‘smoking gun’. In: Key Issues in Environmental Change: Pollution of Lakes and Rivers – a Paleoenvironmental Perspective. MATTHEWS J.A., R.S. BRADLEY, N. ROBERTS and M.A.J. WILLIAMS (Editors), Hodder Arnold, London, pp. 79-94.
- SOMMER U. (1994). The impact of light intensity and day length on silicate and nitrate competition among marine phytoplankton. Limnol. Oceanogr., 39, 1680-1688.
- SPALDING D.J. (1989). The early history of moose (Alces alces): distribution and relative abundance in British Columbia. Contribution to Natural Science, Number 11: 1-2, Royal BC Museum, Victoria, BC.
- STAUFFER R.E. (1990). Granite weathering and the sensitivity of alpine lakes to acid deposition. Limnol. Oceanogr., 35, 1112-1134.
- STEPHENS T., D. ATKIN, U. COCHRAN, P. AUGUSTINUS, M. REID, A. LORREY, P. SHANE and A. STREET-PERROTT (2012). A diatom-inferred record of reduced effective precipitation during the Last Glacial Coldest Phase (28.8–18.0 cal kyr BP) and increasing Holocene seasonality at Lake Pupuke, Auckland, New Zealand. J. Paleolim.,48, 801-817.
- STOERMER E.F. (1977). Postglacial diatom succession in Douglas Lake, Michigan. J. Phycol., 13, 73-80.
- STUIVER M. and P.J. REIMER (1993). Extended 14C database and revised CALIB 3.0 14C age calibration program. Radiocarbon., 35, 215-230.
- TELFORD R.J., H.F. LAMB and M.U. MOHAMMED (1999). Diatom-derived palaeoconductivity estimates for Lake Awassa, Ethiopia: evidence for pulsed inflows of saline groundwater. J. Paleolim., 21, 409-422.
- TELFORD R.J., P. BARKER, S. METCALFE and A. NEWTON (2004). Lacustrine responses to tephra deposition: examples from Mexico. Quat. Sci. Rev., 23, 2337-2353.
- TER BRAAK C.J.F. (1991). CANOCO version 3.12. Agricultural Mathematics Group, University of Wageningen, Wageningen, The Netherlands.
- TER BRAAK C.J.F. and P. ŠMILAUER (1998) Canoco reference manual and user’s guide to Canoco for Windows version 4. Microcomputer Power, Ithaca, NY, USA, 351 p.
- THOMPSON R.I. and K.L. DAUGHTRY (2000). Stratigraphic linkage of carbonate-rich units across east-central Vernon map area, British Columbia: are Kingfisher (Colby) and Big Ledge zinc-lead occurrences part of the same regional marker succession? Current Research 2000-A18, Geological Survey of Canada, pp. 1-5.
- URRUTIA R., A. ARANEDA, F. CRUCES, L. TORRES, L. CHIRINOS, H.C. TREUTLER, N. FAGEL, S. BERTRAND, I. ALVIAL, R. BARRA and E. CHAPRON (2007). Changes in diatom, pollen, and chironomid assemblages in response to a recent volcanic event in Lake Galletue (Chilean Andes). Limnologica, 37, 49-62.
- VERMAIRE J.C., Y.T. PRAIRIE and I. GREGORY-EAVES (2011). The influence of submerged macrophytes on sedimentary diatom assemblages. J. Phycol., 47, 1230-1240.
- VOLLAND S. (2006). Sediment dynamics in Lago Calafquén and Lago Villarrica (Northern Patagonia - Chile): A baseline study on sedimentological processes and considerations in assessing palaeoenvironmental change. PhD Thesis, Technische Universität München, 293 p.
- WALKER I.R. and M.G. PELLATT (2003). Climate change in coastal British Columbia – A paleoenvironmental perspective. Can. Water Resour. J., 28, 531-566.
- WALKER I.R. and M.G. PELLATT (2008). Climate change and ecosystem response in the northern Columbia River Basin - a paleoenvironmental perspective. Environ. Rev., 16, 113-140.
- WECKSTRÖM J., A. KORHOLA and T. BLOM (1997). Diatoms as quantitative indicators of pH and water temperature in sub-arctic Fennoscandian lakes. Hydrobiologia, 347, 171-184.
- WINDER M., J.E. REUTER and S.G. SCHLADOW (2009). Lake warming favours small-sized planktonic diatom species. Proc. Res. Soc. B, 276, 427-435.
- WISSMAR R.C., A.H. DEVOL, J.T. STALEY and J.R. SEDELL (1982). Biological responses on lakes in the Mount St. Helens blast zone. Science, 216, 178-181.
- WOLIN J.A. and H.D. DUTHIE (1999). Diatoms as indicators of water level change in freshwater lakes. In: The Diatoms: Applications for the Environmental and Earth Sciences. STOERMER E.F. and J.P. SMOL (Editors), Cambridge University Press, pp. 183-202.
- WRIGHT H.E., Jr. (1967). A square-rod piston sampler for lake sediments. J. Sed. Petrol., 37, 975-976.
- ZDANOWICZ C.M., G.A. ZIELINSKI and M.S. GERMANI (1999). Mount Mazama eruption: Calendric age verified and atmospheric impact assessed. Geology, 27, 621-624.
- ZIELINSKI G. (2000). Use of paleo-records in determining variability within the volcanism-climate system. Quat. Sci. Rev., 19, 417-438.
 10.7202/033030ar
10.7202/033030arListe des figures
Figure 1
British Columbia with the location of the training set lakes and the Shuswap region (inset). Magnification shows Sicamous Creek Lake (arrow) and the location of other nearby study sites in the ESSF.
La Colombie-Britannique avec la localisation des lacs de la série de calibration ainsi que la région de Shuswap (encadré). Le grossissement montre le lac Sicamous Creek (flèche) et la localisation d'autres sites d'étude à proximité dans l'ESSF.
Figure 2
The diatom stratigraphic record for Sicamous Creek Lake. Taxa with a relative abundance greater than 1% and an occurrence in at least two samples are shown. Abundances are presented as a percentage of total identifiable diatoms.
La stratigraphie des diatomées fossiles du lac Sicamous Creek. Seuls les taxons avec une abondance supérieure à 1 % et qui sont présents dans au moins deux échantillons sont retenus. Les abondances représentent le pourcentage du total des diatomées identifiables.
Figure 3
Benthic versus planktonic diatoms from Sicamous Creek Lake. The uppermost interval (0 cm) is from 1998.
Les diatomées benthiques versus les diatomées planctoniques du lac Sicamous Creek. L'intervalle supérieur (0 cm) date de 1998.
Figure 4
Detrended correspondence analysis (DCA) with four environmental variables, 26 lakes and 110 diatom species (abundance >1% and occurrence in at least two samples) from the British Columbian training set. Numbers represent diatom species (see Table 3), KW-numbers represent lakes, and environmental variable arrows are shown exaggerated (5x).
L'analyse ACD effectuée à partir de quatre variables environnementales, 26 lacs et 110 espèces de diatomées (abondance >1 % et apparition dans au moins deux échantillons) de la série de calibration des lacs de la Colombie-Britannique. Les numéros représentent les espèces de diatomées (voir le Tableau 3), les numéros KW représentent les lacs, et les flèches des variables environnementales ont été exagérées cinq fois.
Figure 5
pH reconstruction with (a) British Columbian transfer function, (b) diversity, (c) evenness and (d) richness in Sicamous Creek Lake.
Les reconstitutions du pH avec (a) la fonction de transfert des lacs de la Colombie-Britannique, (b) la diversité, (c) l’uniformité et (d) la richesse du lac Sicamous Creek.
Figure 6
Correspondence analysis (CA) ordination analysis of fossil diatom abundance showing the ecosystem trajectory, plotted on axes 1 and 2 of the fossil data. Axis 1 shows the sample scores from the first variable (λ = 0.26) and axis 2 gives the sample scores from the second variable (λ = 0.15).
L'ordination de l’analyse de correspondances (AC) de l’abondance des diatomées fossiles montre la trajectoire de l'écosystème, rapporté sur les axes 1 et 2 des données fossiles. L'axe 1 indique les résultats des échantillons pour la première variable (λ = 0.26) et l'axe 2 montre les résultats des échantillons pour la deuxième variable (λ = 0.15).
Figure 7
Correspondence analysis (CA): axis 1 versus axis 2 of the species scores.
L'analyse de correspondances (AC): l'axe 1 versus l'axe 2 des résultats des espèces.
Figure 8
(a) Observed versus inferred WA pH values and (b) plot of residuals.
(a) Valeurs WA déduites versus valeurs WA observées du pH et (b) ordination des résidus.
Liste des tableaux
Table 1
Altitude, physical, chemical, climatic and morphometric variables of the 26 lakes in the British Columbian training set.
L’altitude et les variables physiques, chimiques, climatiques et morphométriques des 26 lacs faisant partie de la série de calibration des lacs de la Colombie-Britannique.
Tableau 2
Correlation matrix of environmental variables, resulting from detrended correspondence analysis (DCA) with the training set data.
La matrice de corrélation des variables environnementales, résultant de l’analyse des correspondances (ACD) avec les valeurs de la série de calibration.
Tableau 3
Numbers for species occurring in detrended correspondence analysis (DCA) plot with training set data.
Les numéros des espèces figurant dans l’analyse ACD faite avec les valeurs de la série de calibration.
Tableau 4
Performance statistics of the British Columbia pH transfer functions.
La performance statistique des modèles de fonction de transfert du pH de la Colombie-Britannique.