Corps de l’article
Les mécanismes moléculaires qui contrôlent la prolifération et la différenciation cellulaires restent à ce jour mal compris. La prolifération et la différenciation sont souvent vues comme des événements antagonistes et de nombreuses études montrent que l’inhibition forcée de la prolifération cellulaire par la surexpression des CKI (cyclin-dependent kinase [CDK] inhibitors) peut induire la différenciation de certains types cellulaires en culture. Par exemple, la surexpression transitoire de l’inhibiteur des CDK p21 (Waf1) dans des cellules leucémiques humaines (lignée U937) entraîne l’expression à la surface cellulaire des marqueurs de différenciation des monocytes et des macrophages CD11b (antigène Mac-1, représentant la chaîne α de l’intégrine hétérodimérique αMβ2) et CD14 (récepteur du lipopolysaccharide) [1]. Dans la majorité des cas, le programme de prolifération et de différenciation des macrophages induit par le M-CSF (macrophage colony-stimulating factor) met en jeu la voie de signalisation passant par Ras qui aboutit à l’activation de facteurs de transcription parmi lesquels on trouve les protéines à domaine Ets tel que Ets-2 [2]. Ces facteurs de transcription se fixent sur deux types de sites: (1) des sites monomériques présents sur le promoteur des gènes impliqués dans le contrôle de la croissance cellulaire comme c-Myc et c-Myb [3, 4]; (2) des sites multimériques AP-1/Ets trouvés dans le promoteur des gènes exprimés lors de la différenciation cellulaire et reconnus par les complexes ternaires Fos/Jun/Ets [5, 6] (Figure 1A). Dans le système hématopoïétique, les facteurs de transcription Ets tels que Ets-1, Ets-2 et Pu-1 jouent un rôle fondamental dans la réponse mitogénique mais aussi dans le programme de différenciation des cellules en réponse aux facteurs de croissance de type CSF (colony-stimulating factors). L’implication des activateurs de transcription à domaine Ets à la fois dans le programme de prolifération et dans celui qui conduit à la différenciation par l’intermédiaire de Ras semble paradoxale. En effet, dans ces cellules, la voie de signalisation de Ras doit être activée pour stimuler l’expression de gènes spécifiques de la différenciation des macrophages mais elle doit, en même temps, être inhibée pour bloquer la réponse proliférative. L’étude réalisée dans le laboratoire de C.K. Glass [7] révèle l’existence d’un nouveau facteur de transcription à domaine Ets, le répresseur Mets (mitogenic Ets transcriptional suppressor) dont la fonction permet de réconcilier les différents effets de la voie Ras sur la prolifération et la différenciation cellulaires.
Figure 1
Mécanisme d’inhibition de la prolifération cellulaire par Mets.
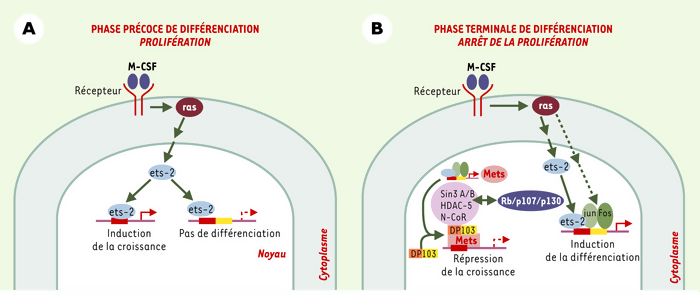
A. Au cours de la prolifération et de la phase précoce de différenciation des macrophages, la voie de signalisation de ras est activée par les facteurs de croissance tel que le M-CSF. Mets n’est pas exprimé et les activateurs de la transcription, tels que Ets-2, se lient à l’ADN et activent principalement la transcription des gènes impliqués dans la croissance cellulaire. B. Au cours de la phase terminale de différenciation, l’expression et l’activation de Ets-2, c-Jun et c-Fos augmentent, induisant l’expression des gènes impliqués dans la maturation des macrophages. Mets se lie spécifiquement sur les sites monomériques et réprime sélectivement la transcription des gènes impliqués dans le contrôle du cycle cellulaire. Cet effet répresseur est réalisé en coopération avec DP103 qui recrute le complexe de répression formé de Sin3 A/B, HDAC-5 et N-CoR, et les protéines de la famille du rétinoblastome (p107, p130, Rb).
Mets a été isolé dans les cellules de la lignée leucémique de monocytes humains THP-1 [7] et dans celles de la lignée d’ostéoblastes murins MC3T3E1 [8]. Au cours de la différenciation des cellules THP-1 en macrophages en réponse à l’ester de phorbol TPA, l’expression de Mets est augmentée [7]. Ce nouveau répresseur possède une forte homologie de séquence avec le facteur de transcription ERF (Ets-2 repressor factor) [9]. Il est composé d’un domaine Ets de liaison à l’ADN et de deux domaines de répression de la transcription en N-terminal et C-terminal. Par son domaine Ets, Mets se lie spécifiquement sur les sites monomériques et réprime ainsi sélectivement les gènes cibles des facteurs Ets, dont on sait qu’ils sont impliqués dans la prolifération dépendante de Ras (Figure 1B). En revanche, il épargne les gènes codant pour des protéines impliquées dans la différenciation, et qui font intervenir Ras. En effet, ces gènes sont contrôlés par les sites multimériques pour lesquels Mets a peu d’affinité. Cette sélectivité permet à Mets, qui est exprimé durant la phase terminale de la différenciation des macrophages, de contribuer à l’arrêt de la croissance cellulaire en inhibant la transcription des gènes impliqués dans la prolifération cellulaire sans affecter les gènes impliqués dans la différenciation. L’action anti-proliférative de Mets est obtenue grâce à l’interaction de Mets avec la protéine DP 103 qui joue le rôle de co-répresseur en recrutant un complexe de répression de la transcription au niveau du promoteur des gènes cibles [7]. DP 103 est une ARN hélicase appartenant à la famille des protéines à domaine DEAD. Cette protéine a été clonée lors de la recherche de partenaires de Mets par des expériences de double-hybride dans la levure [7]. Elle a également été identifiée comme cible de EBNA2 et EBNA3c, protéines impliquées dans l’immortalisation des lymphocytes B par le virus d’Epstein-Barr [10]. Le complexe de répression de la transcription recruté par DP 103 est constitué des histones désacétylases HDAC-2 et HDAC-5, de Sin3 A/B et de N-CoR (Figure 1B). Par ailleurs, l’utilisation de cellules de souris dont les gènes codant pour les protéines de la famille du rétinoblastome (p107, p130 et Rb) ont été inactivés a permis de montrer que ces protéines sont également impliquées dans l’action antiproliférative de Mets [7].
Bien que les mécanismes de régulation de la transcription de Mets restent à definir, une première observation d’ores et déjà remarquable est la présence, dans le promoteur proximal de Mets, de deux sites de fixation du complexe AP-1/Ets. L’expression progressive de Ets-2, c-jun et c-fos au cours de la différenciation des macrophages induit l’expression des gènes spécifiques de la différenciation des macrophages mais pourrait également, par induction de l’expression de Mets, servir de signal pour la sortie du cycle cellulaire.
Parties annexes
Références
- 1. Liu M, Lee MH, Cohen M, Bommakanti M, Freedman LP. Transcriptional activation of the Cdk inhibitor p21 by vitamin D3 leads to the induced differentiation of the myelomonocytic cell line U937. Genes Dev 1996; 10: 142-53.
- 2. Halfon MS, Carmena A, Gisselbrecht S, et al. Ras pathway specificity is determined by the integration of multiple signal-activated and tissue-restricted transcription factors. Cell 2000; 103: 63-74.
- 3. Roussel MF, Davis JN, Cleveland JL, Ghysdael J, Hiebert SW. Dual control of myc expression through a single DNA binding site targeted by ets family proteins and E2F-1. Oncogene 1994; 9: 405-15.
- 4. Sullivan J, Feeley B, Guerra J, Boxer L. Identification of the major positive regulators of c-myb expression in hematopoietic cells of different lineages. J Biol Chem 1997; 272: 1943-9.
- 5. Guidez F, Li AC, Horvai A, Welch JS, Glass CK. Differential utilization of Ras signaling pathways by macrophage colony-stimulating factor (CSF) and granulocyte-macrophage CSF receptors during macrophage differentiation. Mol Cell Biol 1998; 18: 3851-61.
- 6. Karin M, Liu Z, Zandi E. AP-1 function and regulation. Curr Opin Cell Biol 1997; 9: 240-6.
- 7. Klappacher G, Lunyak VV, Sykes DB, et al. An induced Ets repressor complex regulates growth arrest during terminal macrophage differentiation. Cell 2002; 109: 169-80.
- 8. Bidder M, Loewy AP, Latifi T, et al. Ets domain transcription factor PE-1 suppresses human interstitial collagenase promoter activity by antagonizing protein-DNA interactions at a critical AP1 element. Biochemistry 2000; 39: 8917-28.
- 9. Sgouras DN, Athanasiou MA, Beal GJ Jr, Fisher RJ, Blair DG, Mavrothalassitis GJ. ERF: an ETS domain protein with strong transcriptional repressor activity, can suppress ets-associated tumorigenesis and is regulated by phosphorylation during cell cycle and mitogenic stimulation. EMBO J 1995; 14: 4781-93.
- 10. Grundhoff AT, Kremmer E, Tureci O, et al. Characterization of DP103, a novel DEAD box protein that binds to Epstein-Barr nuclear proteins EBNA2 and EBNA3C. J Biol Chem 1999; 274: 19136-44.
Liste des figures
Figure 1
Mécanisme d’inhibition de la prolifération cellulaire par Mets.
A. Au cours de la prolifération et de la phase précoce de différenciation des macrophages, la voie de signalisation de ras est activée par les facteurs de croissance tel que le M-CSF. Mets n’est pas exprimé et les activateurs de la transcription, tels que Ets-2, se lient à l’ADN et activent principalement la transcription des gènes impliqués dans la croissance cellulaire. B. Au cours de la phase terminale de différenciation, l’expression et l’activation de Ets-2, c-Jun et c-Fos augmentent, induisant l’expression des gènes impliqués dans la maturation des macrophages. Mets se lie spécifiquement sur les sites monomériques et réprime sélectivement la transcription des gènes impliqués dans le contrôle du cycle cellulaire. Cet effet répresseur est réalisé en coopération avec DP103 qui recrute le complexe de répression formé de Sin3 A/B, HDAC-5 et N-CoR, et les protéines de la famille du rétinoblastome (p107, p130, Rb).