Corps de l’article
Le streptocoque β-hémolytique du groupe A (Streptococcus pyogenes, SGA) (Figure 1) est un pathogène strictement humain responsable, dans la majorité des cas, d’infections bénignes cutanéo-muqueuses comme l’angine et l’impétigo [1]. Cependant, dans de rares cas, cette bactérie peut être responsable d’infections invasives gravissimes (comme la fasciite nécrosante qui se traduit par la destruction complète des tissus mous) et de syndromes de choc toxique, deux maladies souvent mortelles [2]. En France, des données épidémiologiques récentes montrent que les infections invasives à SGA sont en augmentation depuis 2000, l’incidence des septicémies à SGA étant estimée en 2002 à 1,7 pour 100 000 habitants [3].
Figure 1
Streptocoques du groupe A (Streptococcus pyogenes).
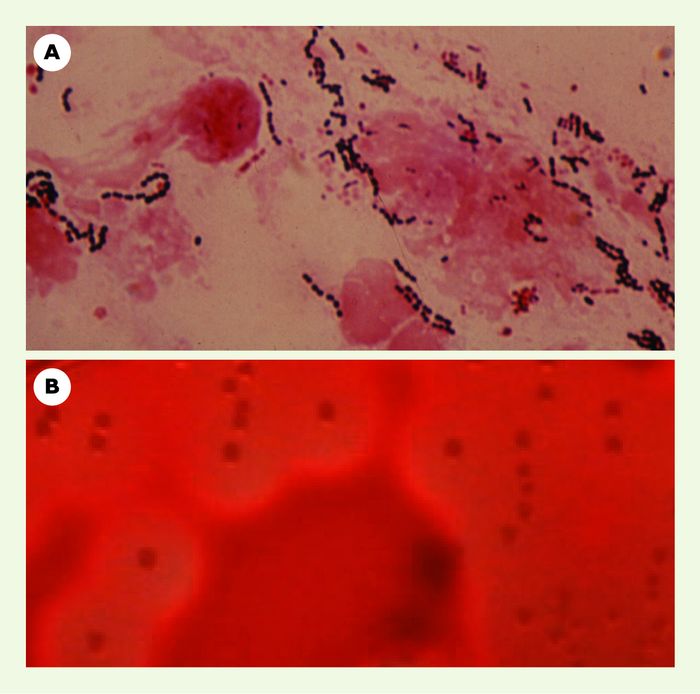
A. Dans un prélèvement de gorge, les streptocoques sont visualisés par la coloration de Gram. B. Culture de 18 heures sur gélose au sang frais. Noter le halo de β-hémolyse autour des colonies.
Le scénario physiopathologique des infections à SGA implique que cette bactérie puisse adhérer et envahir les cellules des muqueuses épithéliales, disséminer dans les tissus sous-jacents et échapper aux défenses immunitaires de l’hôte [1]. Plusieurs déterminants génétiques bactériens impliqués dans le processus physiopathologique des infections à SGA ont été caractérisés [1, 4]. Parmi ceux-ci, la streptokinase (Ska), qui est une sérine protéase, se comporte comme un activateur du plasminogène humain [5]. Chez l’homme, deux types d’activateurs du plasminogène ont été décrits : l’urokinase (uPA) et les activateurs tissulaires dénommés tPA. Ces enzymes, qui sont également des sérine protéases, convertissent le plasminogène en plasmine, enzyme qui ainsi activée facilite la migration des cellules en détruisant les composants de la matrice extracellulaire comme la fibrine, la laminine et la fibronectine. Ces protéases semblent jouer un rôle majeur dans la régulation de la migration des cellules épithéliales, notamment au cours des processus de régénération tissulaire. Ils permettent également la fibrinolyse des caillots sanguins intravasculaires[1] [6]. Le rôle de la streptokinase dans la virulence des SGA avait préalablement été suggéré dans une étude publiée par J. Khil et al. [7]. Dans un modèle d’infection cutanée chez la souris, ces auteurs montraient que la virulence des SGA était augmentée lorsque les bactéries avaient été préalablement incubées en présence de plasma humain ou de plasminogène humain purifié. En revanche, leur virulence (estimée par la taille des nécroses cutanées et la mortalité des animaux) demeurait inchangée lorsque les bactéries avaient été incubées en présence de plasminogène issu d’autres mammifères (rats, lapins, porcs ou chiens). Enfin, l’augmentation de la virulence n’était pas observée pour un mutant bactérien dans lequel le gène ska avait été inactivé. Ces résultats constituaient la première démonstration in vivo du rôle de la streptokinase dans la virulence des SGA et suggéraient que seul le plasminogène d’origine humaine était un substrat pour cette protéase. De plus, il avait été montré que le site de fixation du plasminogène à la surface des SGA était un motif peptidique localisé dans l’une des principales protéines de surface de S. pyogenes, la protéine M codée par les gènes emm [8]. Ces gènes présentent une grande variabilité génétique permettant de définir plus d’une centaine de variants [8]. Seules les souches qui appartiennent au profil D possèdent une séquence peptidique dénommée PAM (plasminogen-binding group A streptococcal M protein) se liant au plasminogène humain avec une forte affinité [9]. Les résultats publiés dans la revue Science par H. Sun et al. [10] démontrent de manière formelle que le plasminogène est un facteur d’hôte primordial contribuant à la virulence des SGA. Les auteurs ont utilisé des souris transgéniques « humanisées » dans lesquelles le transgène codant pour le plasminogène humain sous le contrôle d’un promoteur murin a été introduit chez des souris Plg-/- (Tg-), permettant d’obtenir des concentrations plasmatiques de plasminogène d’une valeur équivalente à 17 % de celles détectées dans le plasma humain. Ces souris, dénommées Tg+ ont été infectées par différentes souches de SGA. Les résultats obtenus ont montré que ces souris Tg+ étaient plus sensibles que les souris témoins Tg- aux infections provoquées par des SGA. Cette hypersensibilité se traduisait par une augmentation de la taille des lésions cutanées observées et du taux de mortalité ; elle est corrélée à l’expression de la streptokinase et nécessite la présence de la séquence PAM à la surface de la bactérie via la protéine M.
Un certain nombre d’hypothèses ont été émises afin d’expliquer comment le recrutement et l’activation du plasminogène par une bactérie pathogène invasive pouvaient contribuer à sa virulence. Il est admis que la réponse inflammatoire induite par un processus infectieux se traduit localement par des thromboses vasculaires et l’occlusion des microvaisseaux. Les résultats publiés par H. Sun et al. [10] suggèrent que les SGA détournent à leur profit le système fibrinolytique de leur hôte pour disséminer dans les tissus avoisinants et la circulation sanguine et échapper ainsi aux défenses locales. Cette hypothèse a également été confirmée dans cette étude puisque le nombre de bactéries retrouvé dans la rate, qui reflète la dissémination hématogène de l’agent infectieux, était plus élevé chez les souris Tg+que chez les souris Tg-. Enfin, les auteurs suggèrent qu’un polymorphisme génétique, responsable de variations des concentrations plasmatiques de plasminogène dans différentes populations humaines, pourrait constituer un facteur de susceptibilité aux infections dues à ces bactéries. L’inféodation des SGA à l’homme apparaît donc être une conséquence de la stricte spécificité de la streptokinase pour le plasminogène humain. Si les résultats décrits dans cet article démontrent de manière irréfutable le rôle de la streptokinase dans la virulence des SGA, il n’en demeure pas moins que d’autres facteurs de virulence contribuent également à la pathogénicité de cette bactérie [4].
Parties annexes
Note
-
[1]
Ces protéases sont d’ailleurs utilisées en thérapeutique humaine comme composés fibrinolytiques dans le cadre de maladies thromboemboliques, telles que l’embolie pulmonaire et l’infarctus du myocarde.
Références
- 1. Cunningham MW. Pathogenesis of group A streptococcal infections. Clin Microbiol Rev 2000 ; 13 : 470-511.
- 2. Bisno AL, Stevens DL. Streptococcal infections of skin and soft tissues. N Engl J Med 1996 ; 334 : 240-5.
- 3. Georges S, Perrocheau A, Laurent E, et al. Infections invasives à Haemophilus influenzae, L. monocytogenes, N. meningitidis, S. pneumoniae, S. agalactiae et S. pyogenes en France en 2000-2002. BEH 2004 ; 34 : 165-8.
- 4. Bisno AL, Brito MO, Collins CM. Molecular basis of group A streptococcal virulence. Lancet Infect Dis 2003 ; 3 : 191-200.
- 5. Lottenberg R, Broder CC, Boyle MD, et al. Cloning, sequence analysis, and expression in Escherichia coli of a streptococcal plasmin receptor. J Bacteriol 1992 ; 174 : 5204-10.
- 6. Kenagy RD, Vergel S, Mattsson E, et al. The role of plasminogen, plasminogen activators, and matrix metalloproteinases in primate arterial smooth muscle cell migration. Arterioscler Thromb Vasc Biol 1996 ; 16 : 373-82.
- 7. Khil J, Im M, Heath A, et al. Plasminogen enhances virulence of group A streptococci by streptokinase-dependent and streptokinase-independent mechanisms. J Infect Dis 2003 ; 188 : 497-505.
- 8. Fischetti V. Surface proteins of Gram positive bacteria. In : Fischetti V et al., eds. Gram positive pathogens. Washington DC : American Society for Microbiology, 2000 : 11-24.
- 9. Svensson MD, Sjobring U, Bessen DE. Selective distribution of a high-affinity plasminogen-binding site among group A streptococci associated with impetigo. Infect Immun 1999 ; 67 : 3915-20.
- 10. Sun H, Ringdahl U, Homeister JW, et al. Plasminogen is a critical host pathogenicity factor for group A streptococcal infection. Science 2004 ; 305 : 1283-6.
Liste des figures
Figure 1
Streptocoques du groupe A (Streptococcus pyogenes).
A. Dans un prélèvement de gorge, les streptocoques sont visualisés par la coloration de Gram. B. Culture de 18 heures sur gélose au sang frais. Noter le halo de β-hémolyse autour des colonies.