Résumés
Abstract
The obliquebanded leafroller (OBLR), Choristoneura rosaceana (Harris) [Lepidoptera: Tortricidae], a primary pest in Quebec apple orchards, can be naturally parasitized. Knowing that habitats around crop’s peripheries are reservoirs for natural enemies of pests, the objective of the present investigation was to assess parasitism and parasitoid guild composition associated with the OBLR. The two-year study included orchards under integrated pest management, their edges, and adjacent woodlands. Parasitism was assessed using sentinel OBLR larvae and considered spring, early summer and late summer. Parasitism rates between regions with different vegetation composition were not significantly different. The first year, late summer larvae showed higher parasitism in orchards (27%), compared to edges (7%) and woodlands (11%). The following year, larvae exposed in early summer had higher parasitism rate in edges (28%) compared to orchards and woodlands (17% in both zones). Nineteen parasitoid species parasitized sentinel larvae. The tachinid Actia interrupta (Curran), the most abundant species, represented 28 and 62% of species the first and the second year respectively. Our research demonstrates that natural biological regulation of the OBLR is the result of a highly diversified parasitoid guild and this should be taken into account in any Integrated Pest Management program.
Keywords:
- parasitism,
- parasitoid diversity,
- Actia interrupta,
- larval parasitoid,
- parasitoid assemblage,
- species richness,
- IPM
Résumé
Choristoneura rosaceana (Harris) [Lepidoptera : Tortricidae] (TBO), ravageur primaire des vergers de pommiers au Québec, peut être naturellement parasitée. Étant donné que les habitats près des périphéries des cultures sont des réservoirs pour les ennemis naturels des ravageurs, l’objectif de cette recherche était d’évaluer le parasitisme et la diversité des parasitoïdes de la TBO. L’étude sur deux ans porte sur les vergers sous régie intégrée, leurs lisières et boisés adjacents. Le parasitisme fut évalué avec des larves sentinelles de TBO, placées à différentes périodes (printemps, début, fin d’été). L’année 1, seules les larves de fin d’été ont présenté des différences significatives entre les zones : parasitisme supérieur en verger (27 %), comparativement à la lisière (7 %) et aux boisés (11 %). L’année suivante, les larves exposées en début d’été présentaient un parasitisme supérieur en lisière (28 %), par rapport aux larves en verger et dans les boisés (17 %). Dix-neuf espèces de parasitoïdes ont attaqué les larves sentinelles de TBO. Le tachinaire Actia interrupta (Curran) était le plus abondant (28 % année 1 et 62 % année 2). Nos résultats démontrent que la régulation naturelle de la TBO est le résultat de l’action d’une guilde très diversifiée de parasitoïdes, qui doit être prise en compte dans tout programme de lutte.
Mots-clés :
- parasitisme,
- diversité de parasitoïdes,
- Actia interrupta,
- parasitoïdes des larves,
- assemblage des parasitoïdes,
- richesse d’espèces,
- lutte intégrée
Corps de l’article
Introduction
The obliquebanded leafroller (OBLR), Choristoneura rosaceana (Harris) (Lepidoptera: Tortricidae), endemic to North America (Beers 1993), is a primary pest in Quebec apple orchards (Morin et al. 2017). However, the OBLR has developed resistance to organophosphate and pyrethroid insecticides in many locations (Carriére et al. 1996). An alternative to chemical approaches is biological control, since natural enemies of leafrollers, particularly parasitoids, can significantly lower pest populations (Pfannenstiel et al. 2012; Unruh et al. 2012). Conservation biological control (CBC) aims to enhance natural enemy efficacy through management of their environment (Eilenberg et al. 2001). Therefore, an assessment and a better understanding of naturally occurring pest enemies in different habitats contribute to improve CBC. A number of studies have demonstrated that habitats along crop’s periphery are key reservoirs for pest’s natural enemies (Gillespie et al. 2016; Jonsson et al. 2008; Wratten et al. 1998). Such habitats are important for parasitoid conservation, since they provide refuges, nutrition sources, and alternative hosts (Gillespie et al. 2016; Lavandero et al. 2006; Letourneau and Altieri 1999; Pfannenstiel et al. 2010, 2012; Unruh et al. 2012). For example, a study showed that flowering plants, planted to attract predators and parasitoids of apple pests in Quebec, increased parasitoid populations by several folds and resulted in an improvement of the quality of the fruit at harvest, which attained 91% in the 5th year of this investigation (Bostanian et al. 2010). Parasitism and parasitoid diversity depends on orchard management and on their adjacent habitats, at a local and at a landscape level (Maalouly et al. 2013) and the persistence of parasitoids can be enhanced by woodland (Boccaccio and Petacchi 2009). Quebec apple orchards are frequently adjacent to woodlands that might harbor natural enemies such as parasitoids, which may also attack apple pests.
The OBLR is bivoltine in southeastern Canada (Hunter and Mcneil 2000) and it overwinters as 3rd instar larvae (Van der Geest and Evenhuis 1991). Overwintering larvae become active in mid-May and remain so until mid-June. A second generation of OBLR follows in early summer, which causes significant damage to apple fruits (Morin et al. 2017). Yet, rates of parasitism of early- and late-summer OBLR larvae in woodlands associated with orchards, along with the parasitoid species involved, has never been assessed. This information is essential to optimize CBC, by managing habitats surrounding orchards in order to promote parasitoid populations.
In Quebec, orchards under Integrated Pest Management (IPM) consider insecticide treatments when needed according to scouting. These can be one application of a pyrethroid (during pink), one application of an organophosphate (petal fall), and one or two applications of organophosphates or reduced risk insecticides. In conventional orchards, applications are not related to scouting and pyrethroids may be used during summer. Treatments of kaolin and microorganisms are used in organic orchards.
Since most orchards in Quebec are under IMP, the aim of the present study was to assess parasitism of OBLR larvae and to characterize the associated parasitoid guild in these orchards and in their adjacent woodlands, by using sentinel OBLR larvae. We hypothesised that parasitism of sentinel OBLR larvae is higher in woodlands than in apple orchards under IPM, due to conditions favoring parasitoids and the absence of chemical interventions in these habitats. Likewise, we expected parasitoid diversity to be higher in woodlands than in orchards.
MATERIALS AND METHODS
Sampling
Sampling was conducted at 6 sites in 2005 (three sites in Montérégie-East and three sites in Brome-Missisquoi) and 10 sites were sampled in 2006 (five sites in Montérégie-East and five sites in Brome-Missisquoi). One experimental unit, consisting of at least 1 ha orchard under IPM, 1 ha of adjacent woodland, and the edge between both, was defined at each site. Sites were selected considering the following: presence of Cortland and Lobo apple cultivars, known preferred host plants by OBLR, standard and semi-dwarf apple trees and previous OBLR infestations.
Sentinel OBLR larvae were used to assess parasitism and parasitoid diversity. The parameters considered were the geographical region (two regions), periods of OBLR larval activity in the orchard (three periods), and zone (three zones: orchard, their edge, and woodland).
Sentinel larvae
Sentinel OBLR larvae were used to assess parasitism. These originated from a rearing at Institut de recherche et de développement en agroenvironnement (IRDA), St-Bruno-de-Montarville, initiated from overwintering larvae collected during the 2004 spring, in an organic apple orchard at Mont Saint-Hilaire (45°31’58.4’’N and 73°9’35.91’’E) and in an experimental orchard at Mont-Saint-Bruno (45°32’33’’N and 73°20’36’’O), Quebec. This rearing was supplemented with new overwintering larvae from the same orchards in 2005. Egg masses ≤ 72 h old were held at 5°C until used (within the following seven days). After hatch, first-instar larvae were individually placed in 5 mL Solo® plastic cups and fed with an artificial diet based on Lima beans (Shorey and Hale 1965). Second instars, as well as fourth instar larvae, were used as sentinel individuals in order to provide younger/smaller and older/larger hosts to parasitoids. Second instar larvae were used in spring, since during the exposition period of time (one week) they could molt to third instar and became acceptable hosts for parasitoids attacking older instars. Fourth instar larvae were exposed late in the summer, as this instar is present in the naturally occurring population, in the case of larvae that haven’t undergone diapause before the critic luminosity threshold (Gangavalli and Aliniazee 1985).
Sentinel larvae were transferred to one-year old apple shoots (10 individuals per apple shoot). These were kept on 1-L water containers at 1.5 m above the ground. The basal portion of shoots was in contact with water and their stems and leaves containing sentinel larvae were exposed, providing easy access for parasitoids to reach sentinel larvae. Each of these apple shoots on water containers was called a “station”.
Region
Plant composition in woodlands adjacent to orchards in each region, was characterized visually using a random systematic design (Barbour et al. 1999). Starting from two random points located on the edge between orchards and woodlands, transects perpendicular to this zone were drawn towards the woodlands. On each transect, two 100 m2 (10 m x 10 m) quadrats were established at 20–30 m and 50–60 m from the edge zone. Species belonging to the Rosaceae family, as well as maple sugar, Acer saccharum (Marsh), were considered OBLR hosts (Chapman and Lienk 1971; Prentice 1965) and the number of their stems was recorded (as the number of stems available for OBLR). The number of conifers and the number of herbaceous plants species in the edge zone were also noted.
Periods of OBLR larval activity
Assessment of parasitism and parasitoid diversity was conducted during three periods of OBLR larval activity in the orchard in order to represent parasitoid species occurring in each of these periods.
Spring = mid-May to mid-June, at the phenological stages of full pink to post-bloom for apple trees.
Early summer = early to late July
Late summer = early August to early September. OBLR larvae active in this period enter diapause and overwinter as immature larvae.
Zone
Three zones or habitats were considered: the orchard, their edge and the adjacent woodland. In 2005, a transect comprising six stations was set up in each of the six sites. One station was placed between the orchard and the woodland, in the edge zone. Two stations were placed in the woodland at 20 m and 40 m distance from the edge, and three stations were placed in the orchard at 20 m, 40 m and 60 m distance from the edge. In 2006 this sampling design was modified for two transects of three stations: one station in the orchard at 40 m, one station in the edge, and one station in the woodland at 40 m. Ten of each, second- and fourth-instar OBRL larvae, were placed and left on each of these stations for one week.
Assessment of parasitism
After being exposed to parasitism, sentinel larvae were individually transferred to 5 mL Solo® cups and fed ad libitum with a Lima bean artificial diet (Shorey and Hale 1965) at 20°C, 16L: 8O, and 60% RH until adulthood, emergence of an adult parasitoid or death.
In addition to sentinel larvae, 54 naturally occurring indigenous overwintering OBLR larvae were collected on May 16th and 17th 2006 in the 11 orchards from the canopy of apple trees located along the perimeter of orchards and also from apple trees located at 15 m from the perimeter. OBLR larvae collected in the orchards were brought to the laboratory and reared under the same conditions as sentinel larvae, i.e., until adulthood, emergence of an adult parasitoid or death.
Parasitoid identification
All parasitoids emerging from sentinel and indigenous OBLRs were preserved in alcohol and identified afterwards using identification keys (Burks 2003; Goulet and Huber 1993; Grissell and Schauff 1990; O’Hara 2005). Identifications were validated by experts from the Canadian National Collection of Insects, Arachnids and Nematodes and from the Natural History Museum of Los Angeles County.
Calculations and statistical analyses
Shannon diversity index (H) (Shannon 1948) for vegetation, the ratio between deciduous trees and conifers (based on basal area), and the proportion OBLR host species (based on the number of available stems in the case of trees and based on total surface covered in the case of herbaceous plants) were calculated for both, woodlands and edges in each of the two regions, Montérégie-East and Brome-Missisquoi. Student’s t-tests were used to compare these variables between both regions.
The proportion of parasitized OBLR larvae was obtained by dividing the number of total parasitized larvae by the total of larvae recovered from the stations and suitable for analyses. Several individuals were unsuitable for analyses as a result of mortality due to causes other than parasitism, which reflect natural mortality excluding parasitism. However, since the number of larvae suitable for analyses was not the same in all sites and stations, the improved angular transformation of Johnson and Kotz (1969) was performed in order to adjust parasitism levels (Sokal and Rohlf 1995). The formula for this adjustment is:
Total parasitism rates were calculated for each region and for each period of OBLR larval activity. Student’s t-tests and the Kruskall-Wallis non-parametric test (SAS institute 2016; Sokal and Rohlf 1995) were used to compare these rates between the regions Montérégie-East and Brome-Missisquoi for each period of OBLR larval activity.
Total parasitism rates in orchards, in edges and in woodlands were calculated for each site and for each period of larval activity. Location relative to the edge zone (distance from the edge), was not considered a variable affecting parasitism or parasitoid diversity in statistical analyses due to the low number of data found in each location point.
Parasitism rates between years were compared by means of a Chi2 test (α = 0.05) and pairwise comparisons, with Bonferroni corrections of the p value performed in order to detect differences among generations of OBLR larvae (Kleinbaum and Klein 2012). Additionally, parasitism rates were compared among the three habitats for each generation of larvae using Chi2 tests. Concerning parasitoid species, small sample sizes did not allow statistical testing to compare distribution among sites or periods of activity of OBLR larvae. However, correlation analyses were performed to examine the relationship between parasitism levels between orchards and woodlands. All statistical analyses were carried out using the JMP® software v.13.1 (SAS Institute, Cary, NC).
RESULTS
Vegetation in each region
Composition of the vegetation in woodlands adjacent to orchards in each region was different. Woodlands in Montérégie-East were entirely composed of deciduous trees and dominated by sugar maple (A. saccharum). Other dominant species were black raspberry (Rubus occidentalis L.) and the big-toothed poplar (Populus grandidentata Michx.). Woodlands in Brome-Missisquoi were dominated by coniferous species, such as Jack pine (Pinus banksiana Lamb.), red pine (Pinus resinosa Ait.), eastern hemlock (Tsuga canadensis L.), white spruce (Picea glauca Moench), and northern white cedar (Thuja occidentalis L.).
There were significantly more host trees for OBLR in Montérégie-East than in Brome-Missisquoi (t(7.53) = 2.448, p = 0.0419) (Table 1).
Table 1
Vegetation characteristics in woodlands and edges adjacent to apple orchards in Montérégie-East and in Brome-Missisquoi

Asterisk indicate statistical significance (*α = 0.05)
Parasitism associated to periods of OBLR larval activity per region
Globally, 15.8 and 14% of OBLR larvae under analyses were parasitized in years 2005 and 2006 respectively (Table 2). Parasitism rates did not show statistically significant differences between regions. In 2005, average parasitism rates for early summer larvae were 10.4 and 15.8% in Montérégie-East and Brome-Missisquoi, respectively (t(1.25) = 0.926, p = 0.499), and 19.7% in Montérégie-East and 14.9% in Brome-Missisquoi for late summer larvae (t(8.46) = -0.178, p = 0.8629). In 2006, spring larvae showed 8 and 1.5% parasitism in Montérégie-East and Brome-Missisquoi respectively (H(1) = 0.383, p = 0.536) and there was 20.7% parasitism in Montérégie-East and 23% in Brome-Missisquoi for early summer larvae (t(36.09) = -0.178, p = 0.597).
Table 2
Sentinel larvae exposed to parasitism in the experimental sites in years 2005–2006 and parasitism rates. Chi2 test (Bonferroni corrected α = 0.008)

Parasitism associated to periods of OBLR larval activity per zone
Comparisons of parasitism per larval activity period, regardless of zones, indicate that in 2006 spring larvae had the lowest OBLR parasitism rate, compared to early and late summer larvae in 2005 and to early summer larvae in 2006 (X2(3) = 108.006, p < 0.0001; Bonferroni corrected α = 0.008) (Table 2). Parasitism levels varied depending on the period of larval activity and the habitat of exposure (Fig. 1). In 2005, parasitism rates of 0 to 20% were found in different sites and no significant differences were detected among orchards (12%), edges (14%) and woodlands (14.5%) in early summer larvae (X2(2) = 0.1749, p = 0.9168). However, late summer larvae showed significantly higher parasitism (27%) in orchards than in woodlands (11%) or in edges (7%) (X2(2) = 16.53, p < 0.001) and it varied from 0 to 54% among sites.
In 2006, parasitism rates of sentinel larvae exposed from mid-May to mid-June (spring), varied from 0 to 13% among sites and there was no significant difference among orchards (0.7%), edges (3.4%) and woodlands (3%) (X2(2) = 3.36, p = 0.1862). However, sentinel larvae from early summer showed higher parasitism in edges (28%) than in orchards or in woodlands (17% in both habitats) (X2(2) = 24.29, p < 0.001). In this case, parasitism rates varied from 0 to 50%.
No significant correlations were found between parasitism in orchards and in woodlands in any of the two years. Early summer larvae in 2005: r = -0.027, n = 5, p = 0.966; late summer larvae in 2005: r = -0.282, n = 12, p = 0.3742; spring larvae in 2006: r = -0.185, n = 39, p = 0.259; early summer larvae in 2006: r = 0.174, n = 39, p = 0.29.
Parasitism of indigenous OBLR larvae
In 2006, 54 naturally occurring overwintering OBLR individuals were found on canopy trees in the 10 apple orchards considered in this study. Parasitism rate was 16.7%.
Figure 1
Percentage of parasitized

sentinel OBLR larvae in 2005 and in 2006, according to period of OBLR larval activity and zone of exposition (Chi2, p < 0.001).
Parasitoid diversity
Regarding diversity of parasitoids, 19 species belonging to 5 families were recovered from sentinel larvae during the two-year study (Table 3). The larval parasitoid, Actia interrupta (Curran) [Diptera: Tachinidae] was the most abundant species both years. This species was responsible for 4.4% of parasitism in 2005 and 8.6% parasitism in 2006. Moreover, A. interrupta represented 28 and 62% of the parasitoid richness found in 2005 and in 2006, respectively. Meteorus trachynotus (Viereck) [Hymenoptera: Braconidae] (4% in 2005 and 20% in 2006) and Hercus fontinalis (Holmgren) (16% in 2005 and 7% in 2006) were moderately present and followed by Oncophanes americanus (Weed) [Hymenoptera: Braconidae], Colpoclypeus florus (Walker) and Sympiesis sp. [Hymenoptera: Eulophidae]. Other species were found occasionally (Fig. 2). Among these, two species were found parasitizing the OBLR for the first time in Quebec, Megaselia longipennis (Malloch) [Diptera: Phoridae] and Tranosemella sp. [Hymenoptera: Ichneumonidae].
Concerning indigenous OBLR larvae, they were parasitized by two braconids, Macrocentrus linearis (Nees) and Apanteles sp., causing 13 and 3.7% parasitism respectively (Table 3).
Table 3
Relative abundance of parasitoid species (%) found parasitizing sentinel OBLR larvae, per period of larval activity in each zone of exposition in 2005 and 2006, and abundance of parasitoid species found in overwintering indigenous OBLR larvae in 2005
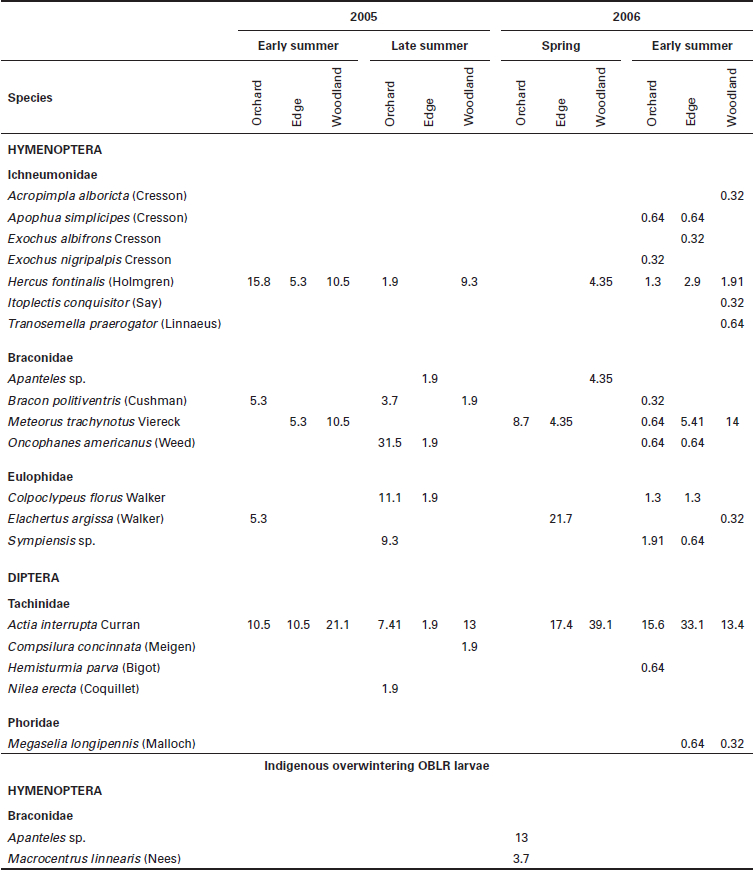
Figure 2
Relative abundance of parasitoid
a)
Year 2005

b)
Year 2006
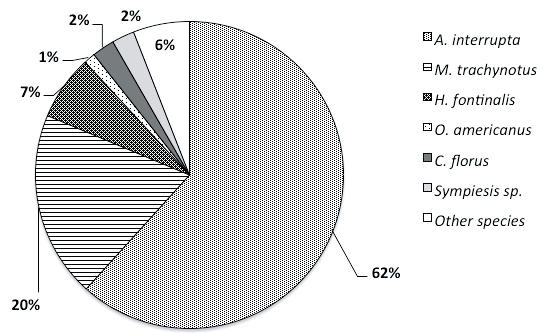
species found in parasitized sentinel OBLR larvae in 2005 (a) and 2006 (b) regardless of OBLR generation and zone of exposition.
DISCUSSION
Parasitism
Unlike as expected, parasitism rates between regions were not different during the two-year study, even though there were more OBLR tree hosts in Montérégie-East than in Brome-Missisquoi. This cannot be explained by temperature differences between regions. Brome-Missisquoi was slightly colder than Montérégie-East in 2005 and 2006 (Tremblay 2008). Other factors that were not examined in the present study might have affected parasitism rates in woodlands. These could be landscape fragmentation (Elzinga et al. 2007; Kruess and Tscharntke 2000) or lack of floral resources for parasitoids in these habitats (Lavandero et al. 2006; Unruh et al. 2012).
Our results do not confirm our original hypothesis, since in general no differences regarding parasitism rates and parasitoid diversity were found between orchards and woodlands. Moreover, no correlation was found between parasitism in orchards and woodlands. Likewise, Sarvary et al. (2010) found that parasitism rates of OBLR in orchards and in adjacent wild habitats were similar, even considering that the latter harbored a higher abundance of leafroller host plants, such as wild apple and wild rose (Rosa spp. L.), which attract alternative hosts for OBLR parasitoids. The authors concluded that resources required by natural enemies were not different in the two habitats. Sarvary et al. (2007b), who obtained similar results of leafroller parasitism in orchards under conventional and reduced-risk insecticide regimes, stated that the combination of mobile natural enemies and habitats treated with toxic insecticides may have masked treatment-specific differences. Likewise, similar parasitism rates in IPM orchards and adjacent woodlands found in the present study could be attributed to a combined effect of analogous suitable resources for parasitoids in both habitats and mobility of these from one habitat to another.
Two exceptions to similar rates of parasitism found among habitats were the higher rate of parasitism in edges as compared to orchards and woodlands for early summer larvae in 2006 and higher parasitism in orchards than in edges and woodlands for late summer larvae of OBLR in 2005. These differences could be due to mobility of parasitoids from woodlands to orchards linked to a higher abundance of parasitoids later in the season, compared to early summer. Moreover, this agrees with the significantly lower parasitism rate found in spring larvae in 2006, compared to parasitism found in early and late summer larvae in 2005 and in early summer larvae in 2006. As other studies have pointed out, abundance of parasitoids tends to be higher in late summer as compared to spring (Cossentine et al. 2004, 2007; Unruh et al. 2012; Wilkinson et al. 2004), mainly due to a higher availability of alternative hosts in this period (Pfannenstiel et al. 2012). In addition, temperatures occurring during mid-May to mid-June are lower than temperatures occurring later in the season, which can also explain differences in parasitoid activity. Mean temperature during May and June were 14 and 18.3°C respectively in sampling sites in 2006, contrasting with 21.4°C in July and 26.7°C in August 2005, and 26.7°C in July 2006 (Tremblay 2008).
Our study shows parasitism rates varying between 0 and 54%, depending on the site and the period of exposure. Besides, there was 15.8% total parasitism in 2005 and 14% total parasitism in 2006, regardless of the zone and periods of OBLR larval activity. Maximum parasitism rate found in this study is higher than parasitism found in some previous studies in North America (Li et al. 1999; Pfannenstiel et al. 2012; Sackett et al. 2007). Conversely, other studies have shown higher parasitism rates than rates found in our investigation (Brunner 1996; Mahr and Whitaker 2004). Such variability in parasitism rates across studies might be representing differences in local influences, such as alternative parasitoid host availability, temperature regimes, and insecticide management in orchards. Likewise, our study shows a wide variability among parasitism rates from one site to another. Yet, it provides useful information on activity of naturally occurring parasitoids species in Quebec.
Parasitoid guild
We recorded 21 species parasitizing the OBLR. From these, two species were associated to indigenous larvae in orchards and a total of 19 were found parasitizing sentinel larvae. Twelve of these were found in orchards, 10 in woodlands and 11 in edges. The diversity of parasitoid species showed in the present investigation is slightly higher than the nine species found in New York State (Sarvary et al. 2007b) and the seven species found in Washington State on sentinel Pandemis pyrusana Kearfott (Lepidoptera: Tortricidae) and OBLR larvae.
Among parasitoid species found in the present study, A. interrupta was the only species consistently present in all sites and generations of OBLR larvae. Hercus fontinalis, M. trachynotus and O. americanus were also widespread but occurred at a lower frequency than A. interrupta. From the 232 tachinid individuals recorded in our study, 228 (98%) were identified as A. interrupta. This is similar to rates found in New York State, where this species represented 94% of tachinids parasitizing the OBLR (Westbrook 2003). Tachinids have previously been mentioned as important parasitoids of leafrollers in several studies (Cossentine et al. 2004; Pfannenstiel et al. 2012; Sarvary et al. 2010; Unruh et al. 2012; Westbrook 2003), and A. interrupta has demonstrated to be one of the more abundant species in several regions. For example, this was the most abundant species in New York State orchards (Sarvary et al. 2007a and b). In another study, the same authors showed that A. interrupta was one of the three most abundant parasitoids recovered from sentinel OBLR larvae (Sarvary et al. 2010) and in Michigan, A. interrupta represented 13% of parasitoid species (Wilkinson et al. 2004). This species is also a main endoparasitoid of the spruce budworm, Choristoneura fumiferana (Lamberth) (Lepidoptera: Tortricidae), one of the most damaging pests of fir and spruce forest in eastern United States and Canada (Cusson et al. 2002). Abundance of this tachinid parasitoid reported in the present as well as in previous research, suggests a competitive advantage of the species. Actia interrupta is ovoviviparous, which means fertilized eggs are incubated and hatch within the reproductive tract of the female. The female larviposits first instar maggots near a host and the maggots search for the host (O’Hara 2005). If the host is already parasitized, and the first parasitoid is still in the egg stage, the maggot can begin feeding immediately, having a size advantage compared to the first parasitoid. Cusson et al. (2002) showed a competitive advantage of A. interrupta over Tranosemea rostrale (Brishke) (Hymenoptera: Ichneumonidae) for parasitism on spruce budworm. The authors concluded that this advantage could be explained by a previous attack of T. rostrale, triggering a reduction in encapsulation by the host. In conclusion, our results, as well as information provided by previous work, show that A. interrupta is a suitable promising species parasitizing the OBLR, and it should be further studied in a conservation biological control perspective.
The parasitoid guild varies depending on OBLR generation. Thus, five parasitoid species were present during mid-May to mid-June, when overwintering OBLR larvae resume diapause and develop to last larval instars: H. fontinalis, Apanteles sp., M. trachynotus, E. argissa et A. interrupta. Identification of indigenous OBLR larvae have confirmed that Apanteles sp. and M. linearis parasitize overwintering larvae. Yet, it was not possible to detect exact time of parasitism occurrence. However, since Apanteles sp. and M. linearis parasitize young larval instars (Cossentine et al. 2005; Krugner et al. 2005), it is likely that parasitism had occurred the previous summer.
Concerning limitations of our study, according to Sarvary et al. (2007a), who used the same methodology to expose sentinel larvae to parasitism, the waterpick method underestimates natural parasitism but it provides an adequate estimation of parasitoid diversity species and demands less resources than using potted apple trees. On the other hand, assessment of natural enemies by using sentinel larvae might bias the results since prey density is abnormal and sentinel larvae behaviour can be unusual. Nevertheless, the present investigation gives valuable information about local occurrence of parasitism on the OBLR, as well as the parasitoid species involved.
Parasitism rates, as well as parasitoid diversity found in this study are relatively similar between habitats. Considering that orchards are habitats disturbed by insecticide treatments, it is likely that woodlands are reservoirs for OBLR parasitoids, which are highly mobile and can fly to orchards quickly after an insecticide treatment. Parasitoid activity should be taken in account to orient insecticide management in orchards and future research. Studies about the impact of recently developed insecticides on natural parasitism and also feasibility of A. interrupta as a biocontrol agent are recommended.
Parties annexes
Acknowledgments
We thank apple growers to allow us access to orchards, Francine Pelletier for technical assistance, Andrew M.R. Bennett, Henri Goulet, John H. Huber, and James E. O’Hara from the Canadian National Collection of Insects, Arachnids, and Nematodes. We also thank Brian V. Brown from the Natural History Museum of Los Angeles County, for confirmation of parasitoid identification. This study was supported by Fonds québécois de la recherche sur la nature et les technologies.
References
- Barbour, M.G., J.H. Burk, W.D. Pitts, F.S. Gilliam, and M.W. Schwartz. 1999. Terrestrial Plant Ecology, Addison Wesley Longman, Menlo Park, California.
- Beers, E.H. 1993. Orchard pest management: a resource book for the Pacific Northwest, Good Fruit Grower, Yakima, WA, USA.
- Boccaccio, L., and R. Petacchi. 2009. Landscape effects on the complex of Bactrocera oleae parasitoids and implications for conservation biological control. Biocontrol 54: 607-616.
- Bostanian, N.J., H. Goulet, J. O’Hara, L. Masner, and G. Racette. 2004. Towards insecticide free apple orchards: flowering plants to attract beneficial arthropods. Biocontrol Sci. Techn. 14: 25-37.
- Brunner, J. 1996. Discovery of Colpoclypeus florus (Walker) [Hymenoptera: Eulophidae] in apple orchards of Washington. Pan-Pac. Entomol. 72: 5-12.
- Burks, R. 2003. Key to the Nearctic genera of Eulophidae, subfamilies Entedoninae, Euderinae, and Eulophinae (Hymenoptera: Chalcidoidea). World Wide Web electronic publication. [http//cache.ucr.edu/%7Eheraty/Eulophidae/] Accessed on May 4, 2006.
- Carriére, Y., J.-P. Deland, and D.A. Roff. 1996.Obliquebanded Leafroller (Lepidoptera: Tortricidae) resistance to insecticides: among-orchard variation and cross-resistance. J. Econ. Entomol. 89: 577-582.
- Chapman, P., and S. Lienk. 1971. Tortricid fauna of apple in New York (Lepidoptera: Tortricidae); including an account of apples’ occurrence in the State, especially as a naturalized plant. New York State Agricultural Experiment Station, Geneva. Pages 87-90.
- Cossentine, J., A. Bennett, H. Goulet, and J. O’Hara. 2007. Parasitism of the spring leafroller (Lepidoptera: Tortricidae) complex in organically managed apple orchards in the north Okanagan valley of British Columbia. Pan-Pac. Entomol. 83: 276-284.
- Cossentine, J., E. Deglow, L. Jensen, and H. Goulet. 2005. Biological assessment of Macrocentrus linearis and Apanteles polychrosidis (Hymenoptera: Braconidae) as parasitoids of the obliquebanded leafroller, Choristoneura rosaceana (Lepidoptera: Tortricidae). Biocontrol Sci. Technol. 15: 711-720.
- Cossentine, J., L. Jensen, E. Deglow, A. Bennett, H. Goulet, J. Huber, and J. O’hara. 2004. The parasitoid complex affecting Choristoneura rosaceana and Pandemis limitata in organically managed apple orchards. BioControl 49: 359-372.
- Cusson, M., M. Laforge, J. Régnière, C. Béliveau, D. Trudel, J. Thireau, G. Bellemare, N. Keirstead, and D. Stolz. 2002. Multiparasitism of Choristoneura fumiferana by the ichneumonid Tranosema rostrale and the tachinid Actia interrupta: occurrence in the field and outcome of competition under laboratory conditions. Entomol. Exp. Appl. 102: 125-133.
- Eilenberg, J., A. Hajek, and C. Lomer. 2001. Suggestions for unifying the terminology in biological control. BioControl 46: 387-400.
- Elzinga, J.A., S. van Nouhuys, D.-J. van Leeuwen, and A. Biere. 2007. Distribution and colonisation ability of three parasitoids and their herbivorous host in a fragmented landscape. Basic Appl. Ecol. 8: 75-88.
- Gangavalli, R., and M. Aliniazee. 1985. Temperature requirements for development of the obliquebanded leafroller, Choristoneura rosaceana (Lepidoptera: Tortricidae). Environ. Entomol. 14: 17-19.
- Gillespie, M.A., G.M. Gurr, and S.D. Wratten. 2016. Beyond nectar provision: the other resource requirements of parasitoid biological control agents. Entomol. Exp. Appl. 159: 207-221.
- Goulet, H., and J.T. Huber. 1993. Hymenoptera of the world: an identification guide to families. Agriculture Canada, Ottawa. 680 pp.
- Grissell, E.E., and M.E. Schauff. 1990. A handbook of the families of Nearctic Chalcidoidea (Hymenoptera). Entomological Society of Washington 1: 1-85.
- Hunter, M.D., and J.N. Mcneil. 2000. Geographic and parental influences on diapause by a polyphagous insect herbivore. Agric. For. Entomol. 2: 49-55.
- Johnson, N.L., and S. Kotz. 1969. Discrete distributions: distributions in statistics. Vol 1. Houghton Mifflin, Boston, USA.
- Jonsson, M., S.D. Wratten, D.A. Landis, and G.M. Gurr. 2008. Recent advances in conservation biological control of arthropods by arthropods. Biol. Control 45: 172-175.
- Kleinbaum, D.G., and M. Klein. 2012. Survival analysis: a self-learning text. Springer-Verlag New York, New York, USA.
- Kruess, A., and T. Tscharntke. 2000. Species richness and parasitism in a fragmented landscape: experiments and field studies with insects on Vicia sepium. Oecologia 122: 129-137.
- Krugner, R., K.M. Daane, A.B. Lawson, and G.Y. Yokota. 2005. Biology of Macrocentrus iridescens (Hymenoptera: Braconidae): a parasitoid of the obliquebanded leafroller (Lepidoptera: Tortricidae). Environ. Entomol. 34: 336-343.
- Lavandero, B., S.D. Wratten, R.K. Didham, and G. Gurr. 2006. Increasing floral diversity for selective enhancement of biological control agents: a double-edged sward? Basic Appl. Ecol. 7: 236-243.
- Letourneau, D. and M.A. Altieri. 1999. Environmental management to enhance biological control in agroecosystems. Pages 319-354 in T.W. Fisher (ed.), Handbook of Biological Control, Principles and Applications of Biological Control. Academic Press.
- Li, S., S. Fitzpatrick, J. Troubridge, M. Sharkey, J. Barron, and J. O’Hara. 1999. Parasitoids reared from the obliquebanded leafroller (Lepidoptera: Tortricidae) infesting raspberries. Can. Entomol. 131: 399-404.
- Maalouly, M., P. Franck, J.-C. Bouvier, J.-F. Toubon, and C. Lavigne. 2013. Codling moth parasitism is affected by semi-natural habitats and agricultural practices at orchard and landscape levels. Agr. Ecosyst. Environ. 169: 33-42.
- Mahr, D., and P. Whitaker. 2004. Beneficial insect habitat in an apple orchard - effects on pests. Available online [https://www.cias.wisc.edu/beneficial-insect-habitat-in-an-apple-orchard%C2%97effects-on-pests/] (Accessed on July 20, 2006).
- Morin, Y., G. Chouinard, and D. Cormier. 2017. La tordeuse à bandes obliques. Fiche 74. Guide de référence en production fruitière intégrée 2016. Réseau-pommier. Available online [http://web2.irda.qc.ca/reseaupommier/?p=10051] (Accessed on March 5, 2018).
- O’Hara, J.E. 2005. A review of the tachinid parasitoids (Diptera: Tachinidae) of Nearctic Choristoneura species (Lepidoptera: Tortricidae), with keys to adults and puparia. Zootaxa 938: 1-46.
- Pfannenstiel, R.S., B.E. Mackey, and T.R. Unruh. 2012. Leafroller parasitism across an orchard landscape in central Washington and effect of neighboring rose habitats on parasitism. Biol. Control 62: 152-161.
- Pfannenstiel, R.S., T.R. Unruh, and J.F. Brunner. 2010. Overwintering hosts for the exotic leafroller parasitoid, Colpoclypeus florus: implications for habitat manipulation to augment biological control of leafrollers in pome fruits. J. Insect Sci. 10: 75.
- Prentice, R.M. 1965. Forest Lepidoptera of Canada recorded by the Forest Insect Survey. Microlepidoptera. Government of Canada, Department of Forestry, Ottawa. Pages 545-834.
- Sackett, T., C. Buddle, and C. Vincent. 2007. Effects of kaolin on the composition of generalist predator assemblages and parasitism of Choristoneura rosaceana (Lep., Tortricidae) in apple orchards. J. Appl. Entomol. 131: 478-485.
- Sarvary, M.A., J. Nyrop, and H. Reissig. 2010. Effects of natural enemies and host plants in wild and orchard habitats on obliquebanded leafroller (Lepidoptera: Tortricidae) larval survival. Biol. Control 55: 110-117.
- Sarvary, M.A., J. Nyrop, and H. Reissig. 2007a. Assessment of three techniques for measuring natural enemy inflicted mortality of leafroller larvae in commercial orchards. Biol. Control 41: 312-320.
- Sarvary, M.A., J. Nyrop, H. Reissig, and K.M. Gifford. 2007b. Potential for conservation biological control of the obliquebanded leafroller (OBLR) Choristoneura rosaceana (Harris) in orchard systems managed with reduced-risk insecticides. Biol. Control 40: 37-47.
- SAS Institute. 2016. JMP® Non parametric. Available on line. [http://www.jmp.com/support/help/Nonparametric.shtml] (Accessed on April 3, 2018).
- Shannon, C.E. 1948. A mathematical theory of communication. Bell Syst. Tech. J. 27: 623-656.
- Shorey, H., and R. Hale. 1965. Mass-rearing of the larvae of nine noctuid species on a simple artificial medium. J. Econ. Entomol. 58: 522-524.
- Sokal, R., and J. Rohlf. 1995. Biometry: the principles and practice of statistics in biological research. W. H. Freeman, New York.
- Tremblay, J. 2008. La tordeuse à bandes obliques dans le sud du Québec : abondance des populations, parasitoïdes associés et influence de divers facteurs biotiques et abiotiques. Master of Biology, Université du Québec à Montréal.
- Unruh, T.R., R.S. Pfannenstiel, C. Peters, J.F. Brunner, and V.P. Jones. 2012. Parasitism of leafrollers in Washington fruit orchards is enhanced by perimeter plantings of rose and strawberry. Biol. Control 62: 162-172.
- Van der Geest, L.P., and H.H. Evenhuis. 1991. Tortricid pests: their biology, natural enemies and control. Elsevier Science Publishers, Bennekom, The Netherlands.
- Westbrook, C.L. 2003. The identity and seasonal activity of tachinid parasitoids of the tortricid leafroller (Choristoneura rosaceana) infesting apple orchards. Master of Science, Cornell University.
- Wilkinson, T.K., D.A. Landis, and L.J. Gut. 2004. Parasitism of obliquebanded leafroller (Lepidoptera: Tortricidae) in commercially managed Michigan apple orchards. J. Econ. Entomol. 97: 1524-1530.
- Wratten, S., H. Van Emden, and M. Thomas. 1998. Within-field and border refugia for the enhancement of natural enemies. Pages 375-403 in C.H. Pickett, and R.L. Bugg (eds.), Enhancing biological control: habitat management to promote natural enemies of agricultural pests. Berkeley.
Liste des figures
Figure 1
Percentage of parasitized
sentinel OBLR larvae in 2005 and in 2006, according to period of OBLR larval activity and zone of exposition (Chi2, p < 0.001).
Liste des tableaux
Table 1
Vegetation characteristics in woodlands and edges adjacent to apple orchards in Montérégie-East and in Brome-Missisquoi
Asterisk indicate statistical significance (*α = 0.05)
Table 2
Sentinel larvae exposed to parasitism in the experimental sites in years 2005–2006 and parasitism rates. Chi2 test (Bonferroni corrected α = 0.008)
Table 3
Relative abundance of parasitoid species (%) found parasitizing sentinel OBLR larvae, per period of larval activity in each zone of exposition in 2005 and 2006, and abundance of parasitoid species found in overwintering indigenous OBLR larvae in 2005