
Résumés
Résumé
La maladie corticale du hêtre (MCH) est de plus en plus fréquente dans les forêts de l’est de l’Amérique du Nord. Des chercheurs ont avancé l’hypothèse que l’expansion de cette maladie pourrait être en partie liée au statut nutritif des arbres. Pour tester cette hypothèse, nous avons vérifié la relation entre l’apparition de cette maladie et la composition du feuillage du hêtre (Fagus grandifolia Ehrh.) dans les stations du Réseau d’étude et de surveillance des écosystèmes forestiers du Québec. Sur les 14 stations contenant du hêtre, cinq présentaient des symptômes de la maladie entre 2006 et 2012. Les arbres infectés étaient en moyenne de plus grand diamètre que ceux qui ne l’étaient pas. La présence de la MCH était liée en partie à la composition foliaire des arbres (pseudo-R2 : 0,551). L’analyse indique que plus le rapport N/P du feuillage du hêtre est élevé, plus la probabilité d’infection par la MCH est importante; et plus les rapports Zn/Mn et Mg/K du feuillage sont élevés, plus la probabilité d’infection par la MCH est faible. Ces rapports donnent à penser que les conditions d’acidification du sol et de drainage influencent la vigueur des arbres et, conséquemment, leur vulnérabilité à la MCH.
Abstract
Beech bark disease (BBD) is becoming more and more frequent in forests of eastern North America. Researchers are bringing forward the hypothesis that the expansion of this disease could partly be related to the trees’ nutrient status. To test this hypothesis, we evaluated the relationship between the occurrence of BBD and the foliage composition of beech (Fagus grandifolia Ehrh.) in plots of the Quebec Forest Ecosystems Study and Monitoring Network. In the 14 plots where beech was present, five displayed BBD symptoms between 2006 and 2012. On average, infected trees had a greater diameter than uninfected trees. The occurrence of BBD was associated in part to tree foliage composition (pseudo-R2: 0.551). The analysis shows that the higher the N/P ratio of the foliage, the higher the probability of infection by BBD; and the higher the Zn/Mn and Mg/K ratios of the foliage, the lower the probability of infection by BBD. These ratios suggest that soil acidification and drainage conditions influence tree vigour and, consequently, its vulnerability to BBD infection.
Corps de l’article
Introduction
Les champignons Neonectria faginata ([Lohman et al.] Castl., Rossman) et Neonectria ditissima ([Tul. et C. Tul.] Samuels et Rossman) sont les agents pathogènes causant la maladie corticale du hêtre (MCH) chez le hêtre à grandes feuilles (Fagus grandifolia Ehrh.) en Amérique du Nord et chez le hêtre européen (Fagus sylvatica L.) en Europe. La germination des spores de ces champignons et leur pénétration dans l’arbre seraient facilitées par les multiples blessures à l’écorce causées par la cochenille du hêtre (Cryptococcus fagisuga Lindinger), un insecte ravageur exotique originaire d’Europe. L’accumulation de chancres sur l’arbre provoque le dépérissement de la cime et, éventuellement, la mort des arbres (Houston 1994).
Au Québec, c’est dans la région du Témiscouata que cette maladie a fait son apparition au début des années 1960. Depuis, la distribution de l’insecte s’étend progressivement vers l’ouest et vers le sud. Les dernières évaluations de la distribution de la maladie au Québec, qui remontent à 2008, indiquent que la maladie était présente dans 90 % des stations d’observation (n = 71) et qu’elle était observable dans la majorité des régions du Québec occupées par le hêtre (MRNF 2009). Les régions les plus atteintes, avec au moins 25 % des arbres atteints par l’un ou l’autre des champignons, étaient celles du Bas-Saint-Laurent, de la Gaspésie–Îles-de-la-Madeleine, de la Capitale-Nationale et de Chaudière-Appalaches. Selon les observations du ministère des Forêts, de la Faune et des Parcs du Québec, la MCH est encore bien présente sur tout le territoire québécois et les dommages sont de plus en plus importants dans plusieurs régions (MRNF 2012). L’absence de patron dans sa distribution spatiale dans le nord-est des États-Unis, où elle est bien installée, donne à penser que cette maladie ne rencontre pas d’obstacles à sa propagation (Garnas et al. 2013).
Des travaux de recherche récents montrent que l’abondance de l’insecte vecteur de la MCH n’est pas le seul facteur prédisposant à cette maladie (Cale et al. 2012; MRNF 2009) et que des facteurs génétiques, environnementaux et physiologiques jouent aussi un rôle. Par exemple, dans les provinces Maritimes de l’est du Canada, la progression de la MCH dans les territoires plus nordiques semble être limitée en partie par les faibles températures hivernales (Taylor et al. 2013). Dans le nord-est des États-Unis, la pente du terrain et la composition du peuplement sont associées à la prévalence de la MCH (McCann et MacDonald 2013). D’autres travaux indiquent que la composition de l’écorce en éléments nutritifs (en particulier, l’azote [N] et le phosphore [P]), en acides aminés et en composés phénoliques est associée à la prévalence de la maladie (Cale et al. 2015). Des chercheurs ont aussi trouvé que la résistance à la MCH avait une base génétique (Mason et al. 2013) et que celle-ci se transmettait aux descendants (Koch et al. 2010). Ainsi, il est possible de réduire l’impact de la MCH par l’aménagement forestier en prélevant les individus affectés et les moins vigoureux (Leak 2006; Taylor et al. 2013).
Étant donné que le hêtre à grandes feuilles prend de l’expansion dans les forêts feuillues du Québec (Duchesne et Ouimet 2008; Duchesne et al. 2005, 2006) et que ce dernier semble de plus en plus affecté par la MCH, il apparaît important de cerner les facteurs qui le prédisposent à cette maladie. C’est dans ce but que nous avons vérifié s’il existe une relation entre la présence de la MCH et la composition foliaire du hêtre dans les stations du Réseau d’étude et de surveillance des écosystèmes forestiers (RESEF) du ministère des Forêts, de la Faune et des Parcs du Québec. La composition foliaire peut nous renseigner sur les conditions de nutrition des arbres qui peuvent être limitantes ou qui, au contraire, peuvent favoriser l’infection des hêtres par la MCH.
Matériel et méthodes
Le RESEF a été mis sur pied en 1986 afin de documenter les impacts des stress environnementaux tels que la pollution atmosphérique, les précipitations acides, les perturbations naturelles et les changements climatiques sur les écosystèmes forestiers. Les stations du réseau sont visitées tous les 5 ans et chaque arbre de diamètre supérieur à 9 cm est examiné pour évaluer son statut (vivant, mort), ses défauts, maladies et caractéristiques dendrométriques (diamètre, hauteur, dimension de la cime). Les stations sont majoritairement composées d’essences feuillues et ont une superficie de 0,5 ha. Le hêtre est présent dans 14 des 43 stations que compte actuellement le réseau (Fig. 1) et la présence de symptômes d’infection par la MCH – présence de laine blanche de cochenilles sur l’écorce, affaissement et fendillement uniforme de l’écorce dans la partie nécrosée qui donne à la tige un aspect verruqueux – a été observée dans cinq de ces stations lors de la dernière visite.
Nous avons relevé les individus dans les stations pour lesquelles nous possédions des données d’analyses foliaires (n = 5 par station). Ces analyses proviennent d’échantillons de feuillage de hêtre prélevés entre le 15 juillet et le 15 août à la dernière visite des stations, soit entre 2006 et 2011. Au total, 45 échantillons foliaires provenant de quatre stations dites saines (stations 402, 702, 703 et 1201) et de cinq stations récemment affectées par la MCH (stations 301, 501, 502, 1202 et 1501) ont fait l’objet d’analyses. Au laboratoire, les échantillons ont été séchés à 65 °C, puis broyés et dissous dans du H2SO4 concentré. Le N (Kjeldahl) a été mesuré par colorimétrie, tandis que les minéraux (P, K, Ca, Mg, Zn et Mn) ont été mesurés par spectrométrie d’émission au plasma. Nous avons ensuite analysé ces données à l’aide d’un modèle de régression logistique avec la présence/absence de MCH comme variable indépendante (transformation logit) et les variables de log-ratio isométriques (LRI) de la composition foliaire en N, P, K, Ca, Mg, Zn et Mn comme variables dépendantes. L’utilisation des valeurs de composition transformées en LRI est en effet nécessaire dans l’analyse de régression afin d’éviter les biais mathématiques, la non-normalité de la distribution et la redondance des variables exprimées en concentration (Parent et al. 2012). Ces valeurs transformées sont en fait des rapports orthogonaux indépendants entre les éléments ou groupe d’éléments. Nous avons employé des rapports entre éléments ayant une interprétation physiologique ou écologique possible selon la littérature (Ågren 2008; Fageria 2001; Parent et al. 2012) (Fig. 2), soit : 1) N-P/Ca-Mg-K-Zn-Mn, 2) N/P, 3) Ca-Mg-K/Zn-Mn, 4) Ca/Mg-K, 5) Mg/K et 6) Zn/Mn. Les données de concentrations foliaires ont été transformées en LRI à l’aide du paquet R compositions (van den Boogaart et al. 2013) et l’analyse de régression logistique a été effectuée dans l’environnement R version 3.0.2 (R Development Core Team 2013).
Figure 1
Stations du réseau RESEF avec présence ou non de la maladie corticale du hêtre (MCH)

Figure 2
Les six rapports indépendants analysés entre les sept éléments du feuillage du hêtre à grandes feuilles

L’échelle représente la proportion de la variance totale de la composition du feuillage.
Résultats et discussion
Évolution de la MCH
Trois stations où la MCH était présente lors de la dernière visite (entre 2006 et 2012; stations 501, 502 et 1202) ont connu une augmentation importante du pourcentage d’arbres atteints par la maladie au cours des cinq années précédentes (Fig. 3). La station 502 est la plus touchée par la maladie avec 50 % des tiges marchandes de hêtre infectées en 2013. Par ailleurs, en 2012, un arbre malade a été répertorié pour la première fois dans la station 1501. La MCH est présente dans la station 301 depuis 1989, mais elle ne s’est pas répandue dans le peuplement. Quant aux stations 105 (qui a été déplacée en l’an 2000), 401 et 1201, des traces de la maladie y ont été notées par le passé, mais elle ne s’y est pas installée définitivement. Aucun signe de la maladie n’a encore été décelé dans les stations 701, 702, 703 et 1502, des stations situées plus à l’ouest dans la province, où l’épidémie est plus récente.
Une portion des arbres montrant des symptômes de la MCH dans les stations du RESEF est morte au cours du suivi (n = 19). Le temps écoulé entre l’observation de l’apparition des symptômes et la mortalité des arbres était de (moyenne ± erreur standard) 5,4 ± 1,2 ans (IC 95 % : 3,0 – 7,9 ans) chez ces arbres. Chez les arbres infectés mais toujours vivants, le temps écoulé depuis l’apparition des symptômes peut aller jusqu’à 8,5 ± 1,1 ans (IC 95 % : 6,4 – 10,6 ans). Cela signifie qu’une certaine proportion des hêtres est capable de tolérer la maladie pendant plusieurs années. Le diamètre des arbres infectés, puis morts, était plus grand d’environ 7 cm que ceux infectés, mais encore vivants (38,5 ± 1,7 cm contre 31,2 ± 2,1 cm; P = 0,011); leur croissance radiale en surface terrière au cours des 10 dernières années était aussi plus faible que celle des arbres infectés, mais toujours vivants (12,8 ± 6,1 cm2 an-1 contre 38,0 ± 5,2 cm2 an-1; P < 0,001).
Figure 3
Évolution de la maladie corticale du hêtre (MCH) d’après les observations effectuées dans 14 stations du RESEF

Les nombres associés à chaque point indiquent le nombre total de hêtres vivants à l’hectare pour l’année d’observation. À noter que la station 105 a été déplacée en 2000.
Figure 4
Résultats de l’analyse de régression logistique
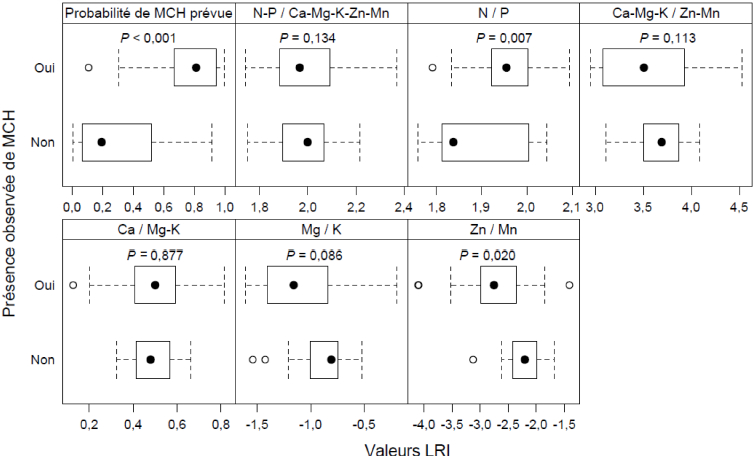
Dans l’ordre habituel (gauche à droite, haut en bas) : présence/absence observée de maladie corticale du hêtre (MCH) dans la station selon la probabilité de présence de MCH prévue par le modèle présenté au tableau 1 : valeurs LRI du rapport N-P/Ca-Mg-K-Zn-Mn; valeurs LRI du rapport N/P; valeurs LRI du rapport Ca-Mg-K/Zn-Mn; valeurs LRI du rapport Ca/Mg-K; valeurs LRI du rapport Mg/K; valeurs LRI du rapport Zn/Mn.
Tableau 1
Coefficients du modèle de régression logistique entre la présence de la maladie corticale du hêtre et la composition foliaire du hêtre

Déviance résiduelle : 37,98; Déviance nulle : 61,83; Pseudo-R2 (de Nagelkerke) du modèle : 0,551.
Les arbres encore vivants infectés par la MCH étaient en général de plus grand diamètre que ceux qui n’étaient pas infectés (31,2 ± 2,1 cm contre 25,9 ± 3,7 cm; P < 0,010) et ils affichaient une croissance radiale en surface terrière au cours des 10 dernières années supérieure à ces derniers (38,0 ± 5,2 cm2 an-1 contre 27,2 ± 2,5 cm2 an-1; P < 0,001). Ainsi, bien que ces arbres conservaient une certaine vigueur, cela n’a pas empêché l’apparition de la MCH.
Composition foliaire
L’analyse de régression logistique montre qu’une relation existe entre la composition foliaire du hêtre et la présence ou absence de MCH dans la station (P < 0,001; Tableau 1). Bien que cette relation n’explique qu’une partie de la variabilité (seulement 55,1 % de la déviance est expliquée par le modèle de régression), elle est de puissance comparable à celles trouvées ailleurs concernant la prévalence de cette maladie (Castello 2014).
L’analyse montre que certains rapports entre des groupes d’éléments du feuillage du hêtre permettent de mieux discriminer la présence/absence de la MCH, en particulier les rapports N/P et Zn/Mn (P ≤ 0,020) et, dans une moindre mesure, le rapport Mg/K (P = 0,086; Fig. 4). Plus le rapport N/P du feuillage du hêtre est élevé, plus la probabilité d’infection par la MCH est grande. Ce résultat va dans le même sens que celui obtenu par l’équipe de Castello qui a trouvé qu’une concentration de l’écorce en P plus faible était associée à la présence de la MCH (Castello 2014; Cale et al. 2015). Le rapport N/P est lié physiologiquement à l’équilibre entre la production des protéines, qui sont riches en N, et la production du catalyseur de la synthèse des protéines – l’acide ribonucléique ribosomique (ARNr) – qui est, pour sa part, riche en P (Loladze et Elser 2011). La baisse du P par rapport à N peut aussi être liée au degré d’acidification du sol. Ce phénomène a pour effet d’augmenter la fixation du P et de réduire son taux de minéralisation par rapport à N qui n’apparaît pas limitatif dans les érablières au Québec (Moore et Houle 2013) en raison des apports atmosphériques importants de cet élément (Ouimet et Duchesne 2009).
Concernant les autres rapports qui semblent associés à la probabilité d’infection par la MCH, plus le rapport Zn/Mn est élevé dans le feuillage, plus la probabilité d’infection par la MCH est faible. Le rapport Zn/Mn est indicateur de drainage, car pour un pH donné, la solubilité du Mn augmente par rapport à celle du Zn à mesure que les conditions de réduction sont favorisées, par exemple lorsque le drainage devient déficient (Bohn et al. 1985). Les hêtres seraient donc plus susceptibles à la MCH dans les stations où le drainage est plus déficient. Ce résultat est en accord avec les travaux de McCann et MacDonald (2013) qui ont observé en Virginie de l’Ouest que la présence de la MCH semblait augmenter lorsque la pente diminuait. Par ailleurs, l’augmentation du Zn dans le feuillage stimule la photosynthèse, particulièrement dans des conditions de sol plus sec (Shkolnik 1984). Finalement, plus le rapport Mg/K est élevé, plus la probabilité d’infection par la MCH est faible. Le rapport Mg/K du feuillage est le reflet du degré de compétition entre ces deux cations pour leur prélèvement (Ouimet et al. 1996) et un indicateur de la production de composés métaboliques par les arbres (Fageria 2001). Une composante physiologique jouerait donc un rôle dans la prévalence de la MCH.
Conclusion
Cette analyse préliminaire a permis de poser des hypothèses permettant d’expliquer en partie la présence de la MCH chez le hêtre à grandes feuilles dans les stations du RESEF. Les résultats donnent à penser que les conditions d’acidification du sol et de drainage peuvent influer sur la vigueur des arbres et, conséquemment, sur leur résistance à la MCH. Les recherches doivent se poursuivre pour valider ces hypothèses.
Parties annexes
Bibliographie
- Ågren, G.I. 2008. Stoichiometry and nutrition of plant growth in natural communities. Ann. Rev. Ecol. Evol. Syst. 39 : 153-170.
- Bohn, H.L., B.L. McNeal et G.A. O’Connor. 1985. Soil chemistry, 2nd ed. John Wiley & Sons, New York.
- Cale, J.A., S.K. Letkowski, S.A. Teale et J.D. Castello. 2012. Beech bark disease: an evaluation of the predisposition hypothesis in an aftermath forest. For. Pathol. 42 : 52-56.
- Cale, J.A., S.A. Teale, M.T. Johnston, G.L. Boyer, K.A. Perri et J.D. Castello. 2015. New ecological and physiological dimensions of beech bark disease development in aftermath forests. For. Ecol. Manag. 336 : 99-108.
- Castello, J.D. 2014. An alternative model of beech bark disease. Final report [En ligne]. http://nsrcforest.org/sites/default/files/uploads/castellocale13full.pdf (consulté le 22 mars 2015).
- Duchesne, L. et R. Ouimet. 2008. Population dynamics of tree species in southern Quebec, Canada: 1970–2005. For. Ecol. Manag. 255 : 3001-3012.
- Duchesne, L., R. Ouimet, J.D. Moore et R. Paquin. 2005. Change in structure and composition of maple-beech stands following sugar maple decline in Québec, Canada. For. Ecol. Manag. 208 : 223-236.
- Duchesne, L., J.D. Moore et R. Ouimet. 2006. Envahissement du hêtre dans les érablières dépérissantes au Québec. Nat. Can. 130 : 56-59.
- Fageria, V.D. 2001. Nutrient interactions in crop plants. J. Plant Nutr. 24 : 1269-1290.
- Garnas, J.R., D.R. Houston, M.J. Twery, M.P. Ayres et C. Evans. 2013. Inferring controls on the epidemiology of beech bark disease from spatial patterning of disease organisms. Agric. For. Entomol. 15 : 146-156.
- Houston, D.R. 1994. Major new tree disease epidemics: Beech bark disease. Ann. Rev. Phytopathol. 32 : 75-87.
- Koch, J.L., D.W. Carey, M.E. Mason et C.D. Nelson. 2010. Assessment of beech scale resistance in full- and half-sibling American beech families. Can. J. For. Res. 40 : 265-272.
- Leak, W.B. 2006. Fifty-year impacts of the beech bark disease in the Bartlett Experimental Forest, New Hampshire. North. J. Am. For. 23 : 141-143.
- Loladze, I. et J.J. Elser. 2011. The origins of the Redfield nitrogen-to-phosphorus ratio are in a homoeostatic protein-to-rRNA ratio. Ecol. Lett. 14 : 244-250.
- Mason, M.E., J.L. Koch, M. Krasowski et J. Loo. 2013. Comparisons of protein profiles of beech bark disease resistant and susceptible American beech (Fagus grandifolia). Prot. Sci. 11 : 1-21.
- McCann, D.P. et W.L. MacDonald. 2013. Preliminary report of ecological factors influencing incidence and severity of beech bark disease in the Appalachian region. Pages 169-181 in G.W. Miller, T.M. Schuler, K.W. Gottschalk, J.R. Brooks, S.T. Grushecky, B.D. Spong et J.S. Rentch (éds.), Proceedings of the 18th Central Hardwood Forest Conference. USDA Forest Service, Northern Research Station, Morgantown, WV. Gen. Tech. Rep. NRS-P-117.
- Moore, J.D. et D. Houle. 2013. Soil and sugar maple response to 8 years of NH4NO3 additions in a base-poor northern hardwood forest. For. Ecol. Manag. 310 : 167-172.
- MRNF. 2009. Insectes, maladies et feux dans les forêts québécoises – 2008. Service des relevés et des diagnostics, Direction de l’environnement et de la protection des forêts, Ministère des Ressources naturelles et de la Faune du Québec.
- MRNF. 2012. Insectes, maladies et feux dans les forêts québécoises – 2011. Service de la gestion des ravageurs forestiers, Direction de la protection des forêts, Ministère des Ressources naturelles et de la Faune du Québec.
- Ouimet, R. et L. Duchesne. 2009. Dépôts atmosphériques dans les forêts au Québec. Retombées actuelles en forêt et tendances au cours des 20 à 30 dernières années. Nat. Can. 133 : 56-64.
- Ouimet, R., C. Camiré et V. Furlan. 1996. Effect of soil K, Ca and Mg saturation and endomycorrhization on growth and nutrient uptake of sugar maple seedlings. Plant Soil 179 : 207-216.
- Parent, S.É., L.E. Parent, D.E. Rozanne, A. Hernandes et W. Natale. 2012. Nutrient balance as paradigm of plant and soil chemometrics. Pages 83-114 in R.N. Issaka (éd.), Soil Fertility [En ligne]. www.intechopen.com/books/soil-fertility (consulté le 22 mars 2015).
- R Development Core Team. 2013. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
- Shkolnik, M.I. 1984. Trace elements in plants. Elsevier Science Publishers, New York.
- Taylor, A.R., D.A. McPhee et J.A. Loo. 2013. Incidence of beech bark disease resistance in the eastern Acadian forest of North America. For. Chron. 89 : 690-695.
- van den Boogaart, K.G., R. Tolosana et M. Bren. 2013. compositions: compositional data analysis. R package version 1.30-2 [En ligne]. http://CRAN.R-project.org/package=compositions (consulté le 22 mars 2015).
Liste des figures
Figure 1
Stations du réseau RESEF avec présence ou non de la maladie corticale du hêtre (MCH)
Figure 2
Les six rapports indépendants analysés entre les sept éléments du feuillage du hêtre à grandes feuilles
L’échelle représente la proportion de la variance totale de la composition du feuillage.
Figure 3
Évolution de la maladie corticale du hêtre (MCH) d’après les observations effectuées dans 14 stations du RESEF
Les nombres associés à chaque point indiquent le nombre total de hêtres vivants à l’hectare pour l’année d’observation. À noter que la station 105 a été déplacée en 2000.
Figure 4
Résultats de l’analyse de régression logistique
Dans l’ordre habituel (gauche à droite, haut en bas) : présence/absence observée de maladie corticale du hêtre (MCH) dans la station selon la probabilité de présence de MCH prévue par le modèle présenté au tableau 1 : valeurs LRI du rapport N-P/Ca-Mg-K-Zn-Mn; valeurs LRI du rapport N/P; valeurs LRI du rapport Ca-Mg-K/Zn-Mn; valeurs LRI du rapport Ca/Mg-K; valeurs LRI du rapport Mg/K; valeurs LRI du rapport Zn/Mn.
Liste des tableaux
Tableau 1
Coefficients du modèle de régression logistique entre la présence de la maladie corticale du hêtre et la composition foliaire du hêtre
Déviance résiduelle : 37,98; Déviance nulle : 61,83; Pseudo-R2 (de Nagelkerke) du modèle : 0,551.