
Résumés
Résumé
Le béluga de l’estuaire du Saint-Laurent (ESL) est une espèce menacée de disparition. L’une des causes contribuant à la précarité de cette population serait liée à son exposition à des composés chimiques toxiques. Cet article de revue collige l’information publiée dans la littérature scientifique sur la contamination des bélugas de l’estuaire du Saint-Laurent par les polluants organiques persistants (POP). Il discute des paramètres et processus qui influencent les concentrations et des tendances temporelles de la contamination observées chez cette population. Enfin, cet article évalue les forces et les limites d’un suivi de la contamination chez le béluga du SL en tant qu’espèce sentinelle de la contamination de son habitat. Entre 1980 et 2007, plus de 20 articles scientifiques ont rapporté la présence de divers POP chez le béluga de l’ESL. Les biphényles polychlorés (BPC) ainsi que le dichlorodiphényltrichloroéthane (DDT) et ses produits de transformation ont été les composés les plus fréquemment rapportés dans les tissus du béluga de l’ESL. La très grande majorité des analyses de contaminants organiques persistants a été réalisée sur des tissus provenant de carcasses de bélugas échouées sur les rives de l’estuaire du Saint-Laurent. Pour les POP dont l’usage est réglementé au Canada, incluant les BPC et le DDT, la contamination du béluga de l’ESL est soit décroissante ou stagnante. Cependant, pour certains composés émergents, notamment les polybromodiphényles éthers, les concentrations doublent à tous les 2-3 ans dans le gras du béluga de l’ESL. Les concentrations et des tendances temporelles de la contamination observées chez cette population sont cependant influencées par plusieurs facteurs dont le sexe et l’âge des animaux ainsi que la période durant laquelle ils ont été exposés aux POP. La qualité et la représentativité des tissus analysés ainsi que certains changements dans les méthodes d’analyses chimiques peuvent affecter les concentrations de POP mesurées chez le béluga. Le programme des carcasses de bélugas de l’ESL constitue un outil précieux pour caractériser la contamination des bélugas par les POP et en évaluer les tendances temporelles. Le béluga de l’ESL ne peut être considéré comme un bon indicateur de la décontamination des POP qui prévalent dans son habitat mais il doit être considéré comme une espèce sentinelle de la présence de POP bioaccumulables en émergence dans son habitat.
Mots clés:
- Contamination,
- mammifère marin,
- béluga,
- Saint-Laurent,
- POP,
- BPC,
- PBDE,
- tendance temporelle
Abstract
The population of beluga from the St. Lawrence Estuary (SLE) is a wildlife species threatened with extinction. One of the possible causes explaining the current status of the population is its exposure to toxic chemicals. This review paper gathers the published data on the contamination of the SLE belugas by persistent organic pollutants (POP). Parameters and processes influencing the concentrations and the temporal trends of POPs in SLE belugas are discussed. The paper also presents the strengths and limits of monitoring contaminants in the SLE belugas for the population itself and as a sentinel species of its habitat. Between 1980 and 2007, more than 20 scientific papers have reported the presence of a variety of POP in the SLE belugas. Polychlorinated biphenyls (PCB) and dichlorodiphenyltrichloroethane (DDT) and its metabolites have been the most frequently reported POP in SLE belugas. Most chemical analyses have been done on beluga tissues collected from stranded carcasses. Concentrations of POP that have been regulated in Canada, including PCB and DDT, are decreasing or stable in SLE belugas. However, concentrations of polybrominated diphenyls ethers, an emerging chemical used as flame retardant, are doubling every 2-3 years in beluga blubber. Concentrations and temporal trends of POP in SLE beluga are influenced by several factors such as the age and sex of animals as well as the time period of their exposure. The quality and the representativeness of samples analyzed or changes in analytical methods can also affect concentrations of POP measured in beluga. In addition, regulations on the use of POP but also changes in beluga diet have affected the exposure of beluga to POP in time. The SLE carcass program has been essential in collecting samples and complementary information on beluga in order to characterize POP contamination and temporal trends in this population. Belugas should not be considered as a good bioindicator species of the decrease of POP in its habitat, but they should be considered as a sentinel species of the presence of bioaccumulative emerging POP entering its habitat.
Keywords:
- Contamination,
- marine mammals,
- beluga,
- St. Lawrence,
- POP,
- PCB,
- PBDE,
- temporal trend
Corps de l’article
1. Introduction
Le béluga (Delphinapterus leucas) de l’estuaire du Saint-Laurent (ESL) est un symbole environnemental au Québec. Cette population réside à l’année dans l’estuaire du Saint-Laurent et fréquente le fjord du Saguenay en saison estivale. Le béluga se trouve à l’origine même de la création du parc marin du Saguenay-Saint-Laurent (PMSSL) en plus de représenter un élément central du projet de la zone de protection marine de l’estuaire du Saint-Laurent (ZPM).
Bien qu’à l’échelle mondiale l’espèce ne soit pas en danger de disparition, il y aurait quelque 300 000 bélugas, la population du béluga de l’ESL possède le statut particulier d’espèce menacée selon le Comité sur la situation des espèces en péril au Canada (COSEPAC). Les principales raisons de ce statut sont, d’une part, l’isolation de cette population par rapport aux populations arctiques et, d’autre part, le manque de rétablissement de cette population décimée par une chasse soutenue entre 1860 et 1960.
D’après HAMMILL et al. (2007), la population du béluga de l’ESL était d’au moins 7 800 individus au milieu du XIXe siècle. En décroissance continue pendant plus de 100 ans, la population aurait été réduite à moins de 1 000 individus à la fin des années 1980. Suite à l’interdiction de la chasse en 1979, une augmentation significative du nombre d’individus était attendue mais le suivi de l’abondance de la population entre 1988 et 2005 montre plutôt que le nombre d’individus est demeuré stable chez cette population. Depuis 1983, un programme des carcasses de bélugas de l’ESL permet de recenser les animaux échoués sur les rives de l’ESL, de récolter des tissus pour différents types d’analyses et, pour les carcasses suffisamment fraîches, de procéder à un examen postmortem afin de déterminer la cause de la mort de l’animal (MEASURES, 2007). En 1995, un plan de rétablissement a été développé pour le béluga de l’ESL, il propose des priorités de recherche et des actions visant à réduire la pression exercée sur le troupeau (BAILEY et ZINGER, 1995).
Plusieurs causes potentielles ont été identifiées pour expliquer le manque de rétablissement de la population du béluga de l’ESL (HAMMILL et al., 2007, MEASURES, 2008). L’exposition des bélugas à des composés chimiques toxiques constitue une des causes pouvant contribuer à la situation précaire de cette espèce dans le Saint-Laurent. Il apparaissait donc essentiel de bien documenter l’exposition de cette population aux composés chimiques toxiques. Les BPC (biphényles polychlorés) et le DDT (dichlorodiphényltrichloroéthane) et ses produits de dégradation ont été les premiers polluants organiques persistants (POP) a être rapportés chez le béluga de l’estuaire du Saint-Laurent. La préoccupation pour les POP réside dans le fait que les concentrations de ces composés augmentent (processus de bioamplification) avec le niveau trophique des organismes dans la chaîne alimentaire aquatique. Le béluga de l’ESL se situe à un niveau trophique élevé parmi les organismes de l’ESL (LESAGE et al., 2001). Par conséquent, les POP sont sujets à atteindre des niveaux élevés chez le béluga de l’ESL, ce qui augmente le risque qu’ils produisent des effets toxiques sur les individus de cette population.
Le but de cet article est de faire un bilan de la contamination du béluga de l’ESL en ciblant les polluants organiques persistants d’origine anthropique. En complément, ce travail identifie les principaux paramètres et processus qui doivent être considérés dans l’interprétation des concentrations et des tendances de la contamination observées chez cette population. Enfin, une discussion portant sur les forces et les limites d’un programme de recherche et de suivi de la contamination chez le béluga de l’ESL complète ce travail.
2. Les polluants organiques persistants mesurés chez le béluga
Les polluants organiques qui s’accumulent dans les organismes possèdent généralement deux caractéristiques fondamentales; ils sont, d’une part, persistants et, d’autre part, lipophiles ou protéinophiles. La persistance d’un composé dans une matrice environnementale est généralement définie en fonction du temps de demi-vie, c’est-à-dire le temps nécessaire pour que la concentration initiale du composé diminue de moitié dans la matrice d’intérêt. Typiquement, un composé est considéré persistant si sa demi-vie est supérieure à deux mois dans l’eau et à six mois dans les sédiments ou les sols (CONVENTION DE STOCKHOLM, 2001, Annexe D). Le caractère lipophile de ces composés, quant à lui, représente la propension d’un composé à se solubiliser dans les lipides. Cette caractéristique est généralement rapportée par une valeur de Kow, c’est-à-dire le rapport de solubilité du composé entre le n-octanol et l’eau. Il a été rapporté que les composés ayant une valeur de Kow entre 104 et 107 sont les plus efficacement bioaccumulés par les organismes et entreposés dans leurs tissus riches en lipides (FISK et al., 1998). Certains composés, généralement des produits de transformation possédant un groupe fonctionnel hydroxyle (OH-) ou méthylsulfonyle (MeSO2-), forment des complexes avec des protéines endogènes favorisant leur rétention et conduisant à leur accumulation dans les organismes (LETCHER et al., 2000a). On retrouve un nombre important de composés chimiques d’origine anthropique qui répondent à ces caractéristiques et que l’on peut mesurer dans les tissus des organismes aquatiques, particulièrement dans le gras des mammifères marins. Parmi ces nombreux composés, on distingue deux groupes principaux, les composés organochlorés et les composés organobromés.
2.1 Les composés organochlorés
Au cours du XXe siècle, de nombreux composés organochlorés ont été synthétisés à des fins industrielles (CAREY et al., 1994). La production de certains de ces composés a conduit à leur émission dans l’environnement ainsi qu’à la formation de sous-produits indésirables. L’amélioration de certains procédés de production a permis de réduire la formation de nombreux produits secondaires. Au cours des dernières décennies, des règlements ont été mis en place afin de bannir l’usage ou de réduire les émissions de certains de ces composés organochlorés dans l’environnement. La convention de Stockholm sur les POP est un accord international élaboré en 2001 puis ratifié en 2004 par plus d’une centaine de pays dont le Canada. Cet accord vise à interdire ou restreindre la production et l’usage de 12 composés organochlorés identifiés comme des polluants organiques persistants prioritaires (CONVENTION DE STOCKHOLM, 2001, Annexe A). En plus des BPC et du DDT, on retrouve dans cette liste les composés suivants : aldrine, chlordane, dieldrine, endrine, heptachlore, hexachlorobenzène, mirex, toxaphène et les polychlorodibenzo-p-dioxines et polychlorodibenzofurannes.
Malgré ces initiatives, plusieurs composés organochlorés et sous-produits d’origine anthropique ou issus de transformations métaboliques continuent d’être bien présents dans les tissus du béluga de l’ESL, notamment dans sa panicule adipeuse ou gras sous-cutané.
2.1.1 Les BPC
Les BPC sont des composés chimiques synthétiques principalement ajoutés aux huiles afin de prévenir leur dégradation et maintenir leur viscosité à des températures élevées. Ils ont été principalement utilisés par l’industrie dans les transformateurs et les condensateurs, les équipements hydrauliques et les lubrifiants. Ils ont aussi été utilisés comme agent plastifiant dans les résines synthétiques. Enfin, les BPC ont été ajoutés à de nombreux produits de consommation courante, notamment dans les peintures, les encres et les cires. La production des BPC a débuté aux États-Unis en 1929, a culminé vers le début des années 1970 pour se terminer en 1979. Sous certaines conditions, notamment à l’intérieur de systèmes clos tels que les transformateurs, l’utilisation des BPC est encore tolérée aujourd’hui. Cependant, aucune nouvelle application des PBC n’a été autorisée au Canada depuis 1983.
Il existe théoriquement 209 congénères de BPC selon la position des atomes de chlore sur les cycles phényles (Figure 1). Les mélanges commerciaux de BPC, connus sous le nom de Aroclor en Amérique du Nord, contiennent un nombre limité de congénères. Les mélanges Aroclor les plus utilisés ont été les Aroclors 1242, 1248 et 1260. Les deux premiers chiffres correspondent au nombre d’atomes de carbone présents sur la molécule, ils sont toujours au nombre de 12 dans les BPC, et les deux derniers chiffres correspondent au pourcentage massique du chlore dans le mélange. Par exemple, 60 % de la masse de l’Aroclor 1260 est constituée de chlore. Les premiers résultats d’analyses quantitatives de BPC faisaient référence aux mélanges commerciaux. Ainsi, la quantification était basée sur un nombre plus ou moins restreint de pics chromatographiques présents dans les échantillons et dans les mélanges Aroclor de référence. Afin d’optimiser la quantification, différents mélanges Aroclor étaient combinés en diverses proportions afin de reproduire le plus possible les patrons de BPC retrouvés dans les échantillons. Les méthodes analytiques courantes sont basées sur la quantification de congénères spécifiques pour lesquels des étalons sont disponibles. Les concentrations de BPC sont généralement rapportées sous la forme de la somme des congénères mesurés (ΣBPC). Il est également assez courant de rapporter les niveaux de BPC par groupes homologues, c’est-à-dire en sommant les concentrations des congénères possédant le même nombre d’atomes de chlore.
Figure 1
Structure chimique générale des biphényles polychlorés (BPC).
General chemical structure of polychlorinated biphenyls (PCB).

La majorité des travaux portant sur la présence de POP chez le béluga de l’ESL rapporte les concentrations de BPC. Les premières données de BPC dans les tissus des bélugas de l’ESL ont été rapportées par SERGEANT (1980). Depuis, 16 études ont rapporté des niveaux de BPC chez le béluga de l’ESL (Tableau 1). Outre l’individu récolté en 1952 (JARMAN et al., 1990), tous les bélugas analysés ont été récoltés entre 1971 et 2003. De plus, l’âge des bélugas analysés couvrent un large spectre, d’un nouveau né (GAUTHIER et al., 1991) jusqu’à un mâle de 74 couches de croissance (CdC) mesurées dans la dentine (LEBEUF et al., 2007a). Les plus récents travaux de STEWART et al. (2007) ont démontré qu’une seule couche de croissance s’accumulait annuellement dans les dents du béluga au lieu des deux couches selon la méthode de calcul utilisée dans les études antérieures. Ainsi, l’animal le plus âgé pour lequel des mesures de contaminants ont été rapportées avait 74 ans. Afin de faciliter les comparaisons, les tableaux 1 et 3 à 7 rapportent les valeurs de CdC après reconversion lorsque les âges étaient rapportés dans les études. Dans le cas des travaux de BENNIE et al. (2000), les valeurs de CdC considérées ont été obtenues à partir des numéros d’identification des animaux et des valeurs de CdC contenues dans la banque de données du ministère des Pêches et Océans (MPO) (L. MEASURES, communication personnelle).
Tableau 1
Concentrations des biphényles polychlorés, BPC, (moyenne ± écart-type, moyenne géométrique et/ou gamme) et contribution relative des groupes homologues (%) dans les tissus des bélugas du Saint-Laurent
Concentrations of polychlorinated biphenyls, PCB, (mean ± std dev, geometric mean and/or range) and relative contribution of homolog groups (%) in tissues of the St. Lawrence belugas
|
|
|
|
|
Moyenne ± écart-type (moyenne géométrique et/ou gamme) |
Contribution des groupes homoloques à la somme arithmétique des BPC ± écart-type (ou à la somme géométrique des BPC) |
|
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
BPC Aroclors (µg•g‑1 lip) |
ΣBPC congénères (µg•g‑1 lip) |
Mono- Tri (%) |
Tétra (%) |
Penta (%) |
Hexa (%) |
Hepta (%) |
Octa-Déca (%) |
Référence |
Gras |
C |
F2 |
(juvénile-adulte) |
1971-1979 |
486 (171-800) |
|
|
|
|
|
|
|
SERGEANT et al., 1980 |
|
C |
M2 |
39 (33-44) |
1983-1984 |
68,4 (68,0) (61,6- 75,2) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
|
C |
M15 |
34 (<1-55) |
1982-1985 |
210 ± 98 (187) (77,7- 344) |
|
|
|
|
|
|
|
MARTINEAU et al., 1987 |
|
C |
F10 |
40 (3,5 -59) |
1982-1985 |
117 ± 211 (50,0) (17,7 – 708) |
|
|
|
|
|
|
|
Idem |
|
C |
M4 |
35 (8-47) |
1986-1987 |
137,1±29,4 (96,3-165,9) |
87,3 ± 17,6 (62,1-103) |
< 0,1 |
18,3 ±1,3 |
11,1 ±1,9 |
36,6 ±8,2 |
26,9 ±8,2 |
7,1 ±3,8 |
MUIR et al., 1990 |
|
C |
F5 |
31 (5-58) |
1986-1987 |
62,6±43,4 (18,9-129,3) |
43,1 ± 25,4 (16,7- 79,3) |
< 0,1 |
15,7 ±12,2 |
14,7 ±6,8 |
34,0 ±22,0 |
27,6 ±16,9 |
8,0 ± 4,8 |
Idem |
|
C |
?3 |
? |
1952- 1986 |
16,6 ± 19,6 (9,5) (2,7-39,0) |
|
|
|
|
|
|
|
JARMAN et al., 1992 |
|
C |
F1 |
50 |
1988 |
|
22,6 |
|
|
|
|
|
|
BERGMAN et al., 1994 |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
(86,0) (7,9-451) |
112,4 ± 98,8 (78,9) (8,3 – 412) |
(< 0,1) |
(13,2) |
(22,7) |
(42,5) |
(18,5) |
(3,2) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987- 1990 |
(30,1) (7,3-90,7) |
35,2 ± 21,0 (29,6) (8,8 – 83,3) |
(0,1) |
(8,5) |
(22,0) |
(39,1) |
(23,9) |
(6,4) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
(97,2) (54,8-145) |
77,3 ± 23,1 (74,5) (48,2 – 126) |
(0,4) |
(9,3) |
(20,4) |
(46,0) |
(19,8) |
(4,1) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
(69,1) (13,5-174) |
77,3 ± 54,9 (57,8) (14,3 – 174) |
(0,3) |
(8,1) |
(20,4) |
(44,2) |
(22,8) |
(4,2) |
Idem |
|
C |
F1 |
68 |
1996 |
|
9,4 |
|
|
|
|
|
|
McALPINE et al., 1999 |
|
C |
?1 |
<1 |
1991 |
|
17,6 |
nd |
13,4 |
17,3 |
50,7 |
17,3 |
1,2 |
GAUTHIER et al., 1998 |
|
B |
M30 |
14* (8-22) |
1994-1996 |
|
7,5 (2,0 - 86,3) |
|
|
|
|
|
|
LETCHER et al., 2000b |
|
B |
F3 |
12* (7-15) |
1994-1996 |
|
10,9 (8,5 – 19,7) |
|
|
|
|
|
|
Idem |
|
B |
M34 |
14* (8-22) |
1994-1998 |
|
20,6 ± 26,8 (12,2) (2,1 – 128) |
(20,3) |
(10,8) |
(19,5) |
(29,0) |
(16,8) |
(3,5) |
HOBBS et al., 2003 |
|
B |
F10 |
14* (7-22) |
1994-1998 |
|
21,2 ± 14,3 (11,8) (0,15- 44,1) |
(8,4) |
(13,2) |
(25,0) |
(38,1) |
(12,7) |
(2,60) |
Idem |
|
C |
M44 |
42 (22-74) |
1988-2002 |
|
85,3 ± 39,5 (75,0) (5,4 – 209) |
0,03 (6,0) |
9,1 (17,9) |
14,8 (19,0) |
49,0 (21,4) |
23,7 (19,9) |
3,3 (15,8) |
LEBEUF et al., 2007a |
|
C |
F42 |
40 (20-64) |
1987-2002 |
|
20,2 ± 17,2 (15,3) (2,6 – 83,1) |
0,13 (7,7) |
6,9 (16,3) |
16,3 (18,7) |
42,4 (20,9) |
28,3 (20,0) |
5,9 (16,4) |
Idem |
Foie |
C |
M3 |
36 (30-44) |
1983-1984 |
5,4 ± 4,9 (4,2) (2,3-11,1) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
|
C |
M10 |
31 (<1-48) |
1983-1985 |
262±153 (233) (61,5-486) |
|
|
|
|
|
|
|
MARTINEAU et al., 1987 |
|
C |
F7 |
37 (3,5 -56) |
1983-1985 |
653±1323 (82,4) (5,5-3869) |
|
|
|
|
|
|
|
Idem |
|
C |
?1 |
<1 |
1991 |
|
8,8 |
|
|
|
|
|
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
53,9 (18,4-81,1) |
0,6 |
12,7 |
18,1 |
46,9 |
14,3 |
7,9 |
METCALFE et al., 1999 |
|
C |
M11 |
21 (<1-48) |
1995-1999 |
|
46,3±39,2 (33,6) (9,3-125) |
|
|
|
|
|
|
DEVIGNE 2003 |
|
C |
F13 |
32(12) (<1-62) |
1995-1999 |
|
39,5±79,5 (12,9) (1,5-293) |
|
|
|
|
|
|
Idem |
|
C |
M3/ F3 |
40 (11-55) 5,3 (<1-9) |
2000-2003 |
|
31,9±6,5 (1,4-45,4) |
< 0,1 |
15,5 |
20,3 |
35,8 |
23,0 |
5,3 |
McKINNEY et al., 2006 |
Rein |
C |
M3 |
36 (30-44) |
1983-1984 |
19 ± 27 (8,6) (3,4-50,2) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
|
C |
M9 |
30 (<1-48) |
1983-1985 |
135±51,4 (124) (57,0-222) |
|
|
|
|
|
|
|
MARTINEAU et al., 1987 |
|
C |
F7 |
37 (3,5 -56) |
1983-1985 |
454±700 (76,2) (4,5-1930) |
|
|
|
|
|
|
|
Idem |
|
C |
?1 |
<1 |
1991 |
|
4,1 |
|
|
|
|
|
|
GAUTHIER et al., 1998 |
Poumon |
C |
M2 |
39 (33-44) |
1983-1984 |
4,4 (4,4) (4,2-4,7) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
Lait |
C |
F1 |
46 |
1984 |
3,8 |
|
|
|
|
|
|
|
MASSE et al., 1986 |
Cerveau |
C |
?1 |
<1 |
1991 |
|
1,7 |
|
|
|
|
|
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
42,9 (8,5-111) |
1,0 |
14,7 |
17,2 |
46,1 |
14,6 |
6,6 |
METCALFE et al., 1999 |
Muscle |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
64,8 (41,2-105) |
0,6 |
13,8 |
15,7 |
48,4 |
14,7 |
6,8 |
METCALFE et al., 1999 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal.
En général, les BPC ont été mesurés dans le gras (sous-cutané) des bélugas, bien que certains travaux aient également rapporté des niveaux de BPC dans d’autres tissus, incluant le foie, le rein, le poumon, le muscle et le lait. Dans presque la totalité des études, les tissus analysés provenaient de carcasses de bélugas échouées sur les rives de l’ESL. Ces tissus étaient récoltés dans le cadre du programme des carcasses de bélugas de l’ESL (MEASURES, 2007). Cependant, deux études, celle de LETCHER et al. (2000b) et celle de HOBBS et al. (2003), ont rapporté des niveaux de BPC dans les gras récoltés à l’aide de la technique par biopsie sur des animaux vivants.
Les niveaux de BPC rapportés dans différents tissus du béluga de l’ESL sont présentés au tableau 1 sous la forme de BPC totaux dont les valeurs ont été estimées en référence à des mélanges Aroclor ou encore sous la forme de la somme de congénères de BPC mesurés. À moins d’indications contraires, les mesures sont rapportées en microgramme (µg) de BPC par gramme de lipide dans le tissu analysé. Lorsqu’il était possible de le faire, la gamme des valeurs était rapportée ainsi que les moyennes arithmétique, avec écart-type, et géométrique. De plus, la contribution des différents groupes homologues, en pourcentage par rapport à la somme des BPC, a été rapportée dans certains travaux.
Les premiers résultats de la contamination du béluga de l’ESL par les BPC ont été rapportés selon la méthode par Aroclor dans du gras provenant de carcasses d’animaux échoués dans les années 1970 (SERGEANT, 1980). Les niveaux rapportés de BPC chez ces animaux sont de l’ordre de la centaine de µg•g‑1 lipide. Les travaux subséquents de MARTINEAU et al. (1987) et MUIR et al. (1990) rapportent aussi, en utilisant la méthode Aroclor, des niveaux du même ordre de grandeur pour les animaux les plus contaminés. Des niveaux de BPC similaires, normalisés sur une base lipidique, ont également été rapportés dans d’autres tissus, notamment le foie et le rein (MARTINEAU et al., 1987).
Les études de MUIR et al. (1990, 1996a et b) rapportent les niveaux de BPC selon les deux méthodes d’analyse. Une comparaison de leurs résultats suggère que les niveaux rapportés par la méthode Aroclor sont légèrement supérieurs à ceux rapportés par la méthode de la somme des congénères. Le tableau 1 rapporte distinctement les niveaux de BPC chez les mâles et les femelles. Certaines études rapportent des niveaux moyens (moyennes géométriques) plus élevés chez les mâles (DEVIGNE, 2003; LEBEUF et al., 2007a; MARTINEAU et al., 1997; MUIR et al., 1990,1996a) alors que d’autres études rapportent des niveaux similaires chez les deux genres (HOBBS et al., 2003; LETCHER et al., 2000b; MUIR et al., 1996b). Les travaux de GAUTHIER et al. (1998) confirment que le transfert des BPC lors de la gestation est important. De plus, la contamination par les BPC dans le gras provenant des biopsies semble plus faible que celle du gras provenant des carcasses, particulièrement chez les mâles. Les résultats de la contribution relative des congénères de BPC des différents groupes homologues dans les tissus des bélugas de l’ESL indiquent une prédominance des congénères ayant entre 4 et 6 atomes de chlore, autant chez les mâles que chez les femelles. Bien qu’une certaine sélectivité dans le transfert des BPC entre la mère et son veau soit attendue, restreignant le transfert des BPC les plus substitués, les patrons des groupes homologues chez les femelles semblent globalement peu affectés par comparaison à ceux des mâles, ce qui est également supporté par un patron semblable chez le nouveau né.
Une comparaison sommaire des niveaux moyens de BPC rapportés dans les gras des bélugas de l’ESL, en considérant l’ensemble des études disponibles (Tableau 1), semble indiquer une tendance à la baisse de la contamination au cours des ans. La première évaluation statistique des tendances temporelles des BPC chez le béluga de l’ESL a été rapportée par MUIR et al. (1996b) en considérant les animaux récoltés entre 1982 et 1994. L’approche utilisée par les auteurs consistait à comparer les concentrations moyennes entre quatre périodes de récolte des animaux, notamment 1982-1885, 1986-1988, 1989-1990 et 1993-1994. Les données pour la période 1982‑1985 provenaient des travaux de MARTINEAU et al. (1987). MUIR et al. (1996b) ont rapporté une diminution des niveaux de BPC, exprimés en équivalent Aroclor, chez les mâles entre 1982-1985 et 1993-1994, alors que chez les femelles, les niveaux ont augmenté au cours de la période examinée. Récemment, LEBEUF et al. (2007a) ont rapporté des diminutions de BPC (somme des congénères) de 3,8 % par année chez les mâles et de 5,1 % chez les femelles. Les tendances temporelles examinées dans cette dernière étude couvraient la période entre 1987 et 2002 et étaient basées sur une approche statistique par régressions linéaires.
2.1.2 Le DDT et ses principaux produits de transformation
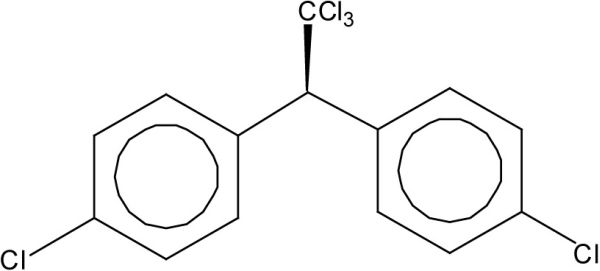
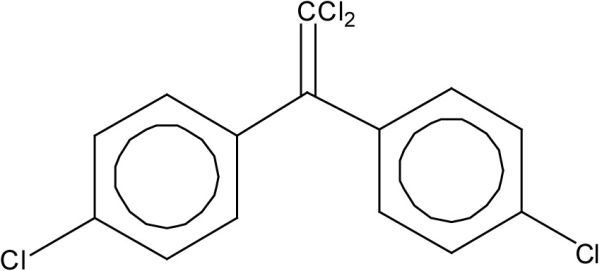
Le DDT a été le principal insecticide utilisé en agriculture en Amérique du Nord entre les années 1940 et 1970. Aux États-Unis, le DDT a été banni en 1973 sauf pour les applications médicales. Au Canada, les principales applications du DDT ont cessé au cours des années 1970 mais le produit n’a été éliminé des registres que vers la fin des années 1980. Le DDT utilisé commercialement est en fait composé à 65-80 % de l’isomère 4, 4’DDT (Figure 2A) et à 15-20 % de l’isomère 2, 4’DDT. Les isomères 4, 4’ et 2, 4’ DDT de ce mélange technique sont transformés par les organismes et dans l’environnement en produits persistants, notamment en 4, 4’DDD, 4, 4’DDE, 2, 4’DDD et 2, 4’DDE, qui s’accumulent eux-mêmes dans les organismes et les sédiments. Parmi les isomères du mélange technique et ses principaux produits de transformation, le 4, 4’DDE est couramment mesuré puisqu’il est généralement le plus abondant dans les organismes (Figure 2B).
Figure 2
Structures chimiques du a) 4, 4’DDT et b) 4, 4’DDE.
Chemical structures of a) 4, 4’DDT and b) 4, 4’DDE.
(a)
(b)
La plupart des études sur la contamination du béluga de l’ESL ont rapporté les niveaux des principaux isomères et produits de transformation du DDT, soit individuellement ou/et sous la forme de la somme des isomères mesurés, ΣDDT (Tableau 2). Il est également courant de rapporter le ratio 4, 4’DDE/ΣDDT. Selon AGUILAR et al. (1987), ce ratio permet d’évaluer si le DDT et ses produits de transformation accumulés dans les organismes proviennent d’une source récente ou ancienne. En effet, le ratio 4, 4’DDE/ΣDDT dans le mélange technique, et par conséquent d’une source récente, est faible, < 0,05, alors qu’il augmente avec le temps en raison de la transformation du DDT en DDE dans l’environnement.
Tableau 2
Concentrations du dichlorodiphényltrichloroéthane, DDT, et ses métabolites (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas du Saint-Laurent*.
Concentrations of dichlorodiphenyltrichloroethane, DDT, and its metabolites (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
Moyenne (µg•g‑1 lipide) ± écart-type (moyenne géométrique et/ou gamme) |
Ratio |
|
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
Sexe, N |
Années |
4, 4’DDT |
4, 4’DDE |
4, 4’DDD |
2, 4’DDT |
2, 4’DDE |
2, 4’DDD |
ΣDDT |
4, 4’DDE /ΣDDT |
Référence |
Gras |
F2 |
1971-1979 |
132 (1,5-262) |
231 (24,9-437) |
(<0,01-128) |
|
|
|
431 (34-827) |
0,63 (0,53-0,73) |
SERGEANT et al., 1980 |
|
M2 |
1983-1984 |
|
81,0 (80,1) (69,0-93,0) |
|
|
|
|
93,7 (92,5) (79,3-108) |
0,87 (0,86) (0,86-0,87) |
MASSE et al., 1986 |
|
M15 |
1982-1985 |
22,4±19,6 (11,0) (0,3-66,1) |
57,9±37,8 (45,9) (8,6-150) |
17,3±10,3 (14,0) (3,2-39,4) |
|
|
|
97,6±64,6 (76,0) (14,5-248) |
0,61±0,07 (0,60) (0,50-0,80) |
MARTINEAU et al., 1987 |
|
F10 |
1982-1985 |
6,8±6,6 (3,9) (0,9-17,3) |
11,4±19,3 (4,2) (1,1-65,1) |
6,1±9,7 (2,6) (0,4-34,2) |
|
|
|
27,2±30,0 (14,8) (2,5-117) |
0,36±0,17 (0,28) (0,05-0,56) |
Idem |
|
M4 |
1986-1987 |
|
75,8±22,0 (44,4-92,9) |
|
|
|
|
116,4±37,6 (60,4-142) |
0,66±0,06 |
MUIR et al., 1990 |
|
F5 |
1986-1987 |
|
16,1±13,0 (2,0-30,9) |
|
|
|
|
26,6±20,0 (4,6-49,3) |
0,56±0,09 |
Idem |
|
?3 |
1952- 1986 |
|
|
|
|
|
|
38,1±39,1 (21,8) (4,4-81,0) |
|
JARMAN et al., 1992 |
|
F1 |
1988 |
|
|
|
|
|
|
16,6 |
|
BERGMAN et al., 1994 |
|
M15 |
1988- 1990 |
(13,6) (0,4-84,3) |
(47,1) (2,1-249) |
(8,6) (0,8-39,2) |
(4,1) (0,05-16,6) |
|
|
120±103 (76,0) (3,4-389) |
0,62 |
MUIR et al., 1996a |
|
F21 |
1987- 1990 |
(3,6) (1,0-15,4) |
(10,4) (1,7-52,3) |
(2,6) (0,6-9,1) |
(0,66) (0,17-3,98) |
|
|
24,3±18,6 (18,6) (3,6-80,8) |
0,56 |
Idem |
|
M9 |
1993-1994 |
(9,3) (1,7-17,8) |
(24,9) (12,6-31,5) |
(6,6) (1,3-11,7) |
(4,4) (0,9-8,5) |
(0,36) (0,16-0,74) |
(0,26) ((0,11-0,45) |
51,5±16,0 (48,4) (20,5-64,6) |
0,51 |
MUIR et al., 1996b |
|
F7 |
1993-1994 |
(7,1) (1,1-20,3) |
(16,6) (3,3-37,5) |
(5,5) (0,9-13,6) |
(2,5) (0,3-9,6) |
(0,23) (0,05-0,51) |
(0,42) (0,29-1,22) |
45,1±27,8 (32,9) (5,7-77,6) |
0,50 |
Idem |
|
F1 |
1996 |
|
|
|
|
|
|
7,3 |
|
McALPINE et al., 1999 |
|
?1 |
1991 |
0,11 |
2,1 |
0,02 |
|
|
|
2,2 |
0,95 |
GAUTHIER et al., 1998 |
|
M30 |
1994-1996 |
|
3,7 (0,3-41,7) |
|
|
0,39 (0,03-4,5) |
|
|
|
LETCHER et al., 2000b |
|
F3 |
1994-1996 |
|
1,5 (0,6-1,8) |
|
|
0,36 (0,3-0,76) |
|
|
|
idem |
|
M5 |
1997-1998 |
|
83,8 (15,5-125) |
|
|
|
|
127 (25,1-190) |
0,66 |
LEBEUF et al., 2001 |
|
F5 |
1997 |
|
6,0 (1,3-9,3) |
|
|
|
|
11,7 (7,1-18,6) |
0,51 |
idem |
|
M34 |
1994-1998 |
(0,94) (0,11-30,2) |
(4,6) (0,4-138) |
(0,97) (0,13-16,9) |
(0,64) (0,07-21,7) |
(0,43) (0,03-4,8) |
(0,39) (<0,01-4,0) |
20,7±36,8 (13,3) (1,3-201) |
0,35 |
HOBBS et al., 2003 |
|
F10 |
1994-1998 |
(0,48) (<0,01-4,6) |
(1,4) (0,05-15,2) |
(0,47) (0,02-2,6) |
(0,23) (<0,01-1,8) |
(0,14) (<0,01-0,8) |
(0,01) (<0,01-4,8) |
7,0±7,3 (5,3) (0,09-24,4) |
0,26 |
idem |
|
M44 |
1988-2002 |
12,6±7,2 (10,6) (0,3-34,4) |
86,9±42,9 (75,8) (2,1-219) |
14,5±7,5 (13,0) (0,7-41,8) |
|
0,30±0,18 (0,25) (0,02-0,81) |
0,31±0,36 (0,25) (0,04-2,5) |
115±55,9 (101) (3,1-297) |
0,76±0,05 (0,75) (0,65-0,88) |
LEBEUF et al., 2007a |
|
F42 |
1987-2002 |
2,2±2,2 (1,4) (0,2-8,9) |
7,1±8,6 (4,3) (0,5-43,0) |
2,2±2,3 (1,4) (0,2-9,6) |
|
0,04±0,06 (0,03) (<0,01-0,4) |
0,08±0,06 (0,06) (0,01-0,3) |
11,6±13,0 (7,3) (0,8-61,2) |
0,60±0,05 (0,59) (0,48-0,71) |
idem |
Foie |
M3 |
1983-1984 |
|
5,4±4,6 (4,0) (1,4-10,5) |
|
|
|
|
5,8±4,8 (4,3) (1,5-11,0) |
0,94±0,02 (0,94) (0,92-0,95) |
MASSE et al., 1986 |
|
M10 |
1983-1985 |
1,9±2,0 (1,4) (<0,01-6,1) |
57,9±36,2 (47,7) (17,5-140) |
32,1±24,8 (23,9) (7,5-85,1) |
|
|
|
91,9±61,7 (74,1) (27,9-232) |
0,65±0,05 (0,64) (0,56-0,71) |
MARTINEAU et al., 1987 |
|
F7 |
1983-1985 |
2,1±1,6 (1,6) (0,3-5,7) |
45,6±101 (6,1) (0,6-293) |
21,4±45,7 (3,7) (0,5-133) |
|
|
|
69,1±148 (12,7) (2,2-432) |
0,52±0,17 (0,48) (0,17-0,70) |
idem |
|
?1 |
1991 |
0,08 |
3,4 |
0,02 |
|
|
|
3,5 |
0,97 |
GAUTHIER et al., 1998 |
|
M4 |
1988-1989 |
5,1 |
48,5 |
9,8 |
|
|
|
63,4 (15,3-75,6) |
0,77 |
METCALFE et al., 1999 |
|
M11 |
1995-1999 |
0,26±0,29 (0,09) (<0,01-0,78) |
24,7±30,4 (12,6) (2,4-94,9) |
6,7±9,2 (3,9) (1,0-32,8) |
|
|
|
31,8±39,1 (17,0) (3,5-128) |
0,74±0,08 (0,74) (0,56-0,87) |
DEVIGNE 2003 |
|
F13 |
1995-1999 |
0,18±0,31 (0,05) (<0,01-1,14) |
22,3±49,0 (3,7) (0,3-163) |
6,1±12,5 (1,4) (0,09-45,5) |
|
|
|
28,6±61,3 (5,4) (0,5-209) |
0,70±0,09 (0,70) (0,53-0,85) |
Idem |
|
M3/ F3 |
2000-2003 |
|
|
|
|
|
|
4,5±1,2 (0,04-7,8) |
|
McKINNEY et al., 2006 |
Rein |
M3 |
1983-1984 |
|
6,7±5,9 (5,1) (2,2-13,4) |
|
|
|
|
7,9±7,9 (5,6) (2,2-16,9) |
0,91±0,10 (0,91) (0,79-0,98) |
MASSE et al., 1986 |
|
M9 |
1983-1985 |
6,0±8,6 (1,7) (0,1-28,0) |
30,8±16,7 (26,4) (11,0-61,6) |
13,4±6,5 (11,4) (3,7-23,0) |
|
|
|
50,2±28,9 (41,9) (18,7-105) |
0,44±0,02 (0,44) (0,41-0,47) |
MARTINEAU et al., 1987 |
|
F7 |
1983-1985 |
3,4±3,2 (2,1) (0,4-10,4) |
40,8±56,9 (8,9) (1,1-147) |
21,0±29,5 (4,9) (0,5-78,9) |
|
|
|
65,3±87,1 (17,0) (2,0-211) |
0,51±0,03 (0,51) (0,45-0,55) |
Idem |
|
?1 |
1991 |
0,04 |
2,3 |
<0,01 |
|
|
|
2,3 |
0,98 |
GAUTHIER et al., 1998 |
Poumon |
M2 |
1983-1984 |
|
6,3 (6,2) (6,0-6,5) |
|
|
|
|
6,6 (6,6) (6,4-6,8) |
0,95 (0,94) (0,94-0,95) |
MASSE et al., 1986 |
Lait |
F1 |
1984 |
|
4,2 |
|
|
|
|
4,5 |
0,92 |
MASSE et al., 1986 |
Cerveau |
?1 |
1991 |
0,01 |
0,69 |
<0,01 |
|
|
|
0,70 |
0,98 |
GAUTHIER et al., 1998 |
|
M4 |
1988-1989 |
27,0 |
86,9 |
6,6 |
|
|
|
121 (9,3-387) |
0,72 |
METCALFE et al., 1999 |
Muscle |
M4 |
1988-1989 |
10,3 |
68,2 |
15,7 |
|
|
|
94,2 (53,7-150) |
0,72 |
METCALFE et al., 1999 |
nombre,
mâle,
femelle,
information sur l’état et l’âge des animaux au Tableau 1
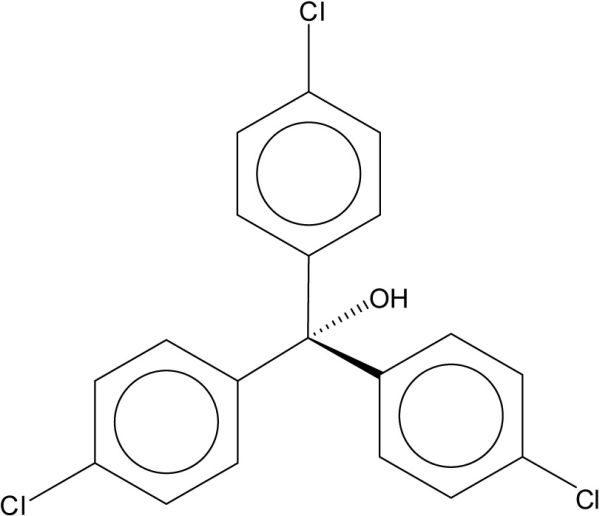
Comme c’est le cas pour les BPC, les isomères du DDT et leurs produits de transformation sont présents à des niveaux mesurables dans différents tissus du béluga de l’ESL. Le 4, 4’DDE représente généralement une grande proportion de la ΣDDT. Puisque le nombre de composés constituant la ΣDDT varie selon les études, il est souvent plus simple de comparer les niveaux du 4, 4’DDE. La gamme de niveaux moyens (moyennes géométriques) rapportée varie généralement entre 10 et 100 µg de 4, 4’DDE par gramme de lipide. Cependant, les niveaux dans les échantillons de gras provenant des biopsies (HOBBS et al., 2003; LETCHER et al., 2000) semblent sensiblement plus bas que ceux provenant des carcasses de bélugas. La contamination des mâles semblent aussi systématiquement plus élevée que celle des femelles. Les ratios 4, 4’DDE/ΣDDT sont difficiles à comparer entre les études en raison du nombre variable de constituants considéré dans la ΣDDT. On note cependant que les ratios varient principalement entre 0,5 et 0,8, ce qui indique que la source de DDT dans le milieu n’est pas récente. Deux situations extrêmes sont également observées. Les ratios 4, 4’DDE/ΣDDT dans les tissus du nouveau né sont très élevés (GAUTHIER et al., 1998) suggérant un transfert préférentiel du 4, 4’DDE par rapport aux autres composés de la ΣDDT, ce qui est supporté par des ratios généralement plus faibles chez les femelles que chez les mâles. La seconde situation extrême concerne les faibles ratios dans les échantillons provenant des biopsies (HOBBS et al., 2003).
Sur la base des niveaux du 4, 4’DDE et/ou de la ΣDDT dans le gras des bélugas rapportés par les 14 études identifiées au tableau 2, aucune diminution de la contamination au cours des ans ne semble apparente chez les mâles ou les femelles. MUIR et al. (1996b) ont cependant rapporté une augmentation significative des niveaux de DDE et ΣDDT chez les femelles entre 1982-1985 et 1993-1994. Chez les mâles, une diminution a été rapportée pour le DDE au cours de cette période alors que la ΣDDT n’aurait diminué qu’à partir de 1986-1988 (MUIR et al., 1996b). Enfin, LEBEUF et al. (2007a) ont rapporté des diminutions de la ΣDDT de 3,7 % par année chez les mâles et de 7,0 % chez les femelles pour la période entre 1987 et 2002. Cependant, la tendance temporelle du 4, 4’DDE n’était pas significative chez les mâles alors qu’une diminution des concentrations était observée chez les femelles.
2.1.3 Le toxaphène
Le toxaphène est un mélange complexe composé principalement de polychlorobornanes et qui contient au moins 1 000 composés. Il a été principalement utilisé comme insecticide dans le nord du Mexique et dans les états du sud des États-Unis. Le Canada aurait utilisé entre 1 et 4 % de la production totale de toxaphène. Cette production a débuté en 1946 pour se terminer en 1981. L’utilisation la plus intense du toxaphène a eu lieu au milieu des années 1970 à la suite du bannissement du DDT.
À l’instar des BPC, le toxaphène a d’abord été quantifié en référence aux principaux pics chromatographiques contenus à la fois dans le mélange technique et les échantillons environnementaux. Puisque le toxaphène technique subit une importante dégradation dans l’environnement, un nombre limité de polychloroboranes s’accumule par les organismes, notamment chez les mammifères marins. La plupart des études qui ont rapporté des niveaux de toxaphène total dans les bélugas de l’ESL réfèrent à la quantification d’environ 8 pics chromatographiques généralement observés (MUIR et al., 1990). Au cours des années, la quantification de congénères spécifiques du toxaphène est devenue de plus en plus courante mais limitée à un nombre restreint de polychlorobornanes, notamment les P26 et P50 selon la notation de Parlar (Figure 3) ou respectivement T2 et T12 selon l’identification utilisée par MUIR et al. (1996a, b).
Figure 3
Structures chimiques des congénères de toxaphène a) P26 et b) P50.
Chemical structures of toxaphene congeners a) P26 and b) P50.
(a)

(b)

Le toxaphène n’a été rapporté que dans un seul tissu du béluga, le gras, récolté soit sur des animaux vivants à l’aide de la technique par biopsie, soit sur des carcasses échouées. Les résultats de toxaphène total rapportés par MUIR et al. (1990, 1996 a,b) sur des carcasses semblent plus élevés que ceux de HOBBs et al. (2003) chez des animaux vivants échantillonnés au cours d’une période plus récente (Tableau 3). Les niveaux des congénères spécifiques P26 et P50 rapportés par MUIR et al. (1996a, b) et GOUTEUX et al. (2003) sont très similaires. En général, les résultats des concentrations de toxaphène total indiquent que cette contamination chez le béluga est plus faible que celle par les BPC ou la ΣDDT. Comme dans la plupart des études rapportant les niveaux de BPC et de la ΣDDT, les mâles sont plus contaminés que les femelles.
Tableau 3
Concentrations du toxaphène total et de certains congénères spécifiques (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans le gras des bélugas du Saint-Laurent.
Concentrations of total toxaphene and selected congeners (mean ± std dev, geometric mean and/or range) in blubber of the St. Lawrence belugas.
|
|
|
|
|
Moyenne (µg•g‑1 lipide) ± écart-type (moyenne géométrique et/ou gamme) |
|
|||
|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
Toxaphène total |
Somme de six congénères |
P26 (T2) |
P50 (T12) |
Référence |
Gras |
C |
M4 |
35 (8-47) |
1986-1987 |
16.9±2.8 (13.5-20.3) |
|
|
|
MUIR et al., 1990 |
|
C |
F5 |
31 (5-58) |
1986-1987 |
7.3±4.1 (2.4-11.9) |
|
|
|
Idem |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
(18.0) (2.4-45.6) |
|
(1,3) (0,20-4,3) |
(4,3) (0,64-16,6) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987- 1990 |
(8.6) (3.2-2.3) |
|
(0,45) (0,17-6,1) |
(1,5) (0,37-22,3) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
(27.0) (16.9-35.9) |
|
(0,80) (0,49-1,1) |
(1,6) (1,1-2,2) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
(20.8) (6.3-40.0) |
|
(0,50) (0,12-1,0) |
(1,3) (0,73-2,4) |
Idem |
|
B |
M30 |
14* (8-22) |
1994-1996 |
(8.0) (1.0-89.7) |
|
|
|
HOBBS et al., 2003 |
|
B |
F5 |
14* (7-22) |
1994-1996 |
(2.4) (0.28-6.5) |
|
|
|
Idem |
|
C |
M26 |
41 (22-58) |
1988-1999 |
|
2,6±0,8 (1,0-4,9) |
0,80±0,24 (0,35-1,3) |
1,7±0,6 (0,61-3,5) |
GOUTEUX et al., 2003 |
|
C |
F26 |
42 (20-63) |
1987-1999 |
|
1,0±0,8 (0,22-3,3) |
0,31±0,27 (0,05-1,2) |
0,57±0,48 (0,16-2,0) |
Idem |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal
Les travaux de MUIR et al. (1996b) indiquent que les concentrations de toxaphène total ont augmenté chez les mâles et les femelles entre 1986 et 1994. Ces observations contrastent avec celles de GOUTEUX et al. (2003) qui rapportent une tendance significative à la baisse pour la majorité des congénères spécifiques examinés chez les femelles et dans une moindre proportion chez les mâles entre 1988 et 1999. En combinant les données de MUIR et al. (1996b) à celles de GOUTEUX et al. (2003) pour les congénères P26 et P50, ces derniers ont rapporté une diminution des concentrations de ces deux congénères chez les femelles alors qu’elles sont demeurées constantes chez les mâles.
2.1.4 Le chlordane, le hexachlorocyclohexane, hexachlorobenzène, Mirex et TCPM
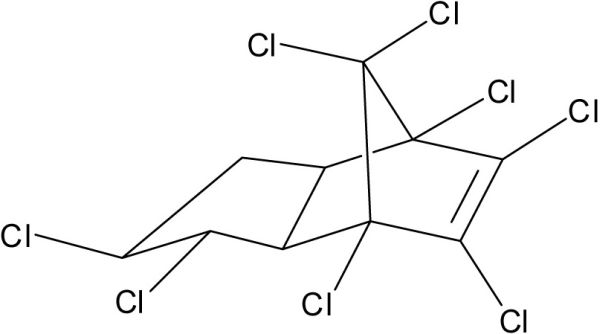
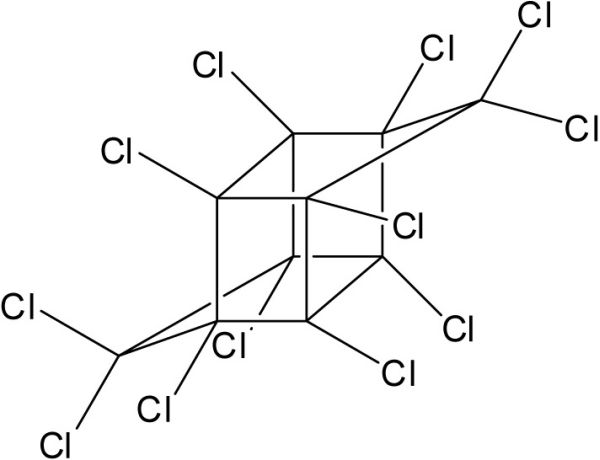
Le chlordane technique (CHL) n’est pas un composé unique mais plutôt un mélange de nombreux produits chimiques apparentés incluant des isomères de chlordanes (Figure 4A), heptachlores, octachlores et nonachlores. Il a été utilisé comme insecticide aux États-Unis et au Canada dès 1949. L’usage du CHL a été progressivement restreint à partir de 1983 puis interdit au début des années 1990.
Figure 4
Structures chimiques du a) γ-Chlordane, b) Lindane (γ-HCH), c) hexachlorobenzène (HCB), d) Mirex et e) tris-(4-chlorophényle) méthanol (TCPMOH).
Chemical structures of a) γ-Chlordane, b) Lindane (γ-HCH), c) hexachlorobenzene (HCB), d) Mirex and e) tris-(4‑chlorophenyl) methanol (TCPMOH).
(a)
(b)

(c)

(d)
(e)
Selon les différentes études, le nombre de composés du CHL mesurés dans les tissus du béluga varie entre quatre (LEBEUF et al. 2007a) jusqu’à onze composés, dont certaines structures restent à être élucidées (HOBBS et al., 2003; MUIR et al., 1990, 1996a,b). Malgré cela, les concentrations de CHL rapportées dans les tissus des bélugas varient peu, entre 1 et 10 µg•g‑1 lipide, suggérant que les principaux composés du CHL ont été considérés dans ces études (Tableau 4). Par comparaison, les niveaux de CHL rapportés dans le rein et le cerveau du nouveau-né analysé par GAUTHIER et al. (1998) sont plus faibles. Par ailleurs, il semble que les concentrations de CHL chez les femelles soient systématiquement plus faibles que chez les mâles lorsque les niveaux de contamination des deux genres sont disponibles pour comparaison et proviennent de la même étude.
Tableau 4
Concentrations du chlordane, hexachlorocyclohexane, hexachlorobenzène, mirex et tris(4-chlorophényle) méthane/méthanol (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas du Saint-Laurent.
Concentrations of chlordane, hexachlorocyclohexane, hexachlorbenzene, mirex and tris(4-chlorophenyl) methane/methanol (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (moyenne géométrique et/ou gamme) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
CHL (µg.g‑1 lip) |
HCH (µg•g‑1 lip) |
HCB (µg•g‑1 lip) |
Mirex (µg•g‑1 lip) |
TCPM (ng•g‑1 lip) |
Référence |
Gras |
C |
M4 |
35 (8-47) |
1986-1987 |
8,6±0,7 (7,5-9,2) |
0,43±0,13 (0,32-0,59) |
1,5±0,50 (0,94-2,2) |
1,2±0,74 (0,22-1,8) |
|
MUIR et al., 1990* |
|
C |
F5 |
31 (5-58) |
1986-1987 |
4,1±2,3 (1,7-7,4) |
0,28±0,12 (0,14-0,37) |
0,69±0,50 (0,25-1,5) |
1,3±0,11 (0,44-3,1) |
|
Idem |
|
C |
?3 |
nd |
1952- 1986 |
|
|
|
|
15±22 (1-41) |
JARMAN et al., 1992 |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
(8,4) (1,4-28) |
(0,47) (0,20-0,90) |
(1,4) (0,24-5,5) |
(0,33) (<0,01-6,8) |
(434) (45-1783) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987- 1990 |
(3,5) (1,2-10) |
(0,29) (0,11-0,77) |
(0,37) (0,08-1,8) |
(1,1) (0,18-5,0) |
(274) (111-701) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
(11) (8,2-16) |
(0,35) (0,28-0,51) |
(0,97) (0,48-1,9) |
(1,0) (0,66-1,9) |
(409) (220-1044) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
(8,9) (2,3-18) |
(0,30) (0,18-0,56) |
(0,67) (0,11-1,7) |
(0,90) (0,18-2,4) |
(336) (220-832) |
Idem |
|
C |
?1 |
<1 |
1991 |
1,4 |
0,16 |
0,28 |
0,06 |
|
GAUTHIER et al., 1998 |
|
C |
M5 |
38 (24-54) |
1997-1998 |
|
|
|
|
34 (8,8-50) |
LEBEUF et al., 2001 |
|
C |
F5 |
57 (42-62) |
1997 |
|
|
|
|
13 (3,5-18) |
Idem |
|
B |
M30 |
14* (8-22) |
1994-1998 |
(2,0) (0,25-8,1) |
(0,53) (0,05-3,6) |
(0,18) (0,04-0,85) |
(0,12) (0,02-1,6) |
(3,1) (2,1-9,0) |
HOBBS et al., 2003 |
|
B |
F5 |
14* (7-22) |
1994-1998 |
(1,4) (0,11-3,4) |
(0,30) (0,05-1,8) |
(0,13) (0,01-1,1) |
(0,14) (<0,01-0,97) |
(4,2) (2,6-6,1) |
Idem |
|
C |
M44 |
42 (22-74) |
1988-2002 |
4,6±1,6 (4,3) (0,78-9,5) |
0,06±0,04 (0,05) (0,02-0,27) |
0,69±0,38 (0,60) (0,13-2,1) |
1,0±0,66 (0,74) (0,04-3,0) |
117±50 (109) (<0,01-260) |
LEBEUF et al., 2007a |
|
C |
F42 |
40 (20-64) |
1987-2002 |
1,3±0,76 (1,1) (0,32-3,2) |
0,08±0,05 (0,06) (0,01-0,22) |
0,20±0,27 (0,13) (0,03-1,5) |
0,71±0,52 (0,50) (0,01-2,4) |
20±18 (15) (<0,01-90) |
Idem |
Foie |
C |
?1 |
<1 |
1991 |
1,5 |
0,24 |
0,67 |
0,10 |
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
2,2 |
0,22 (0,21-0,24) |
2,0 (0,42-4,0) |
0,81 |
|
METCALFE et al., 1999 |
|
C |
M8 |
28 (16-48) |
1995-1999 |
|
|
0,62 (0,19-1,3) |
1,2 (0,08-2,5) |
|
DEVIGNE 2003 |
|
C |
F10 |
38 (10-62) |
1995-1999 |
|
|
0,58 (0,05-3,6) |
0,66 (0,05-2,4) |
|
DEVIGNE 2003 |
|
C |
M3/ F3 |
40 (11-55)/ 5,3 (<1-9) |
2000-2003 |
2,6 ±0,61 (0,06-4,2) |
0,07±0,02 (<0,01-0,14) |
0,12±0,02 (0,02-0,17) |
0,05±0,01 (<0,01-0,08) |
|
McKINNEY et al., 2006 |
Rein |
C |
?1 |
<1 |
1991 |
0,81 |
0,18 |
0,44 |
0,06 |
|
GAUTHIER et al., 1998 |
Cerveau |
C |
?1 |
<1 |
1991 |
0,20 |
0,21 |
0,16 |
0,03 |
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
1,2 |
0,42 (0,29-0,55) |
1,4 (0,28-3,4) |
0,57 |
|
METCALFE et al., 1999 |
Muscle |
C |
M4 |
35(3) (28-40) |
1988-1989 |
4,7 |
0,18 (0,09-0,27) |
2,6 (1,0-6,2) |
1,1 |
|
METCALFE et al., 1999 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal
Selon MUIR et al. (1996b), les niveaux du CHL dans le gras des bélugas étaient à la hausse entre 1986 et 1994 chez les femelles alors qu’ils étaient stables chez les mâles. Ces résultats sont en partie semblables à ceux de LEBEUF et al. (2007a) qui ne rapportent aucune tendance significative du CHL chez les bélugas adultes récoltés entre 1987 et 2002. Cependant, cette dernière étude rapporte des diminutions significatives des isomères alpha (cis) et gamma (trans) chlordanes chez les mâles au cours de cette période.
Le HCH (hexachlorocyclohexane) technique contient plusieurs isomères dont les alpha, bêta, gamma et thêta dans des proportions respectives d’environ 64 %, 10 %, 13 % et 9 %. Le lindane est l’isomère gamma (γ-HCH) et l’ingrédient insecticide actif (Figure 4B). Le mélange HCH a été produit et utilisé comme insecticide aux États-Unis à partir de 1945. Depuis la fin des années 1970, les usages agricoles du HCH ont été progressivement restreints puis interdits aux États-Unis et au Canada mais le lindane, isolé des autres HCH, continue d’être utilisé comme médicament. Le lindane et les autres isomères du HCH sont considérés toxiques pour de nombreux organismes incluant les mammifères.
La plupart des études ont rapporté les niveaux des trois principaux isomères du HCH dans les tissus du béluga à l’exception de l’étude de LEBEUF et al. (2007a) qui n’a retenu que les isomères alpha et gamma alors que celles de GAUTHIER et al. (1998) et de METCALFE et al. (1998) ont considéré les quatre isomères. Ces différences pourraient expliquer les niveaux de HCH sensiblement plus faibles rapportés par LEBEUF et al. (2007a) dans le gras des bélugas bien que ces derniers soient similaires à ceux rapportés par McKINNEY et al. (2006) dans les foies (Tableau 4). Les isomères, alpha, bêta et gamma contribuent dans des proportions similaires à la ΣHCH dans les différents tissus du béluga à l’exception du cerveau où plus de 80 % du HCH est représenté par l’isomère alpha (GAUTHIER et al., 1998, METCALFE et al., 1998).
Le HCH a significativement diminué dans le gras des bélugas mâles entre 1986 et 1994 selon MUIR et al. (1996b) et entre 1988 et 2002 selon LEBEUF et al. (2007a), cette dernière étude rapportant un taux de diminution de 7,5 % par année. Chez les femelles, seule l’étude de LEBEUF et al. (2007a) rapporte une tendance temporelle significative caractérisée par une diminution de 9,4 % par année. Les diminutions importantes du HCH chez le béluga, principalement de l’isomère alpha, pourrait expliquer en partie les faibles niveaux moyens rapportés par LEBEUF et al. (2007a) compte tenu que cette étude, ainsi que celle de McKINNEY et al. (2006), considèrent des animaux récoltés récemment.
L’hexachlorobenzène (HCB) a été utilisé principalement comme fongicide entre 1945 et la fin des années 1960 aux États-Unis et au Canada (Figure 4C). Le HCB a aussi été produit involontairement lors de la fabrication de plusieurs composés organochlorés. Cette source a cependant diminué avec l’amélioration des technologies de fabrication, le contrôle accru des rejets ou l’arrêt de la fabrication de certains composés organochlorés. Le HCB fait partie de la famille des chlorobenzènes (CB) qui compte plusieurs autres produits industriels, incluant certains tétra et penta-CB considérés toxiques et réglementés ou restreints au Canada et aux États-Unis.
Le HCB a été rapporté dans les tissus des bélugas à des concentrations variant entre 0,2 et 2 µg•g‑1 lipide (Tableau 4). MUIR et al. (1996a, b) ont conclu que le HCB était le principal composé de la famille des CB à s’accumuler chez le béluga en mesurant que de faibles niveaux d’isomères des penta et tétra-CB. Cependant, HOBBS et al. (2003) ont rapporté des niveaux de tétra-CB du même ordre que ceux du HCB, soit environ 0,2 µg•g‑1 lipide, alors que les niveaux des di- et tri-CB étaient nettement plus élevés, variant entre 4 et 8 µg•g‑1 lipide.
Des diminutions significatives du HCB chez les mâles et des niveaux stables chez les femelles ont été rapportés par MUIR et al. (1996b) pour la période 1986-1994. LEBEUF et al. (2007a) arrivent aux mêmes conclusions pour la période 1987-2002 et caractérisent la diminution chez les mâles par un taux de 6,9 % par année.
Le Mirex est une molécule complexe et fortement chlorée (Figure 4D). L’utilisation du Mirex comme insecticide a débuté dans les années 1960 aux États-Unis puis a cessé vers la fin des années 1970. Le Mirex n’aurait jamais été utilisé à cette fin au Canada. Le Mirex est aussi un ignifugeant ajouté à de nombreux matériaux. Cette application du Mirex est acceptée aux États-Unis mais elle est interdite au Canada.
Les niveaux moyens de Mirex sont d’environ 1 µg•g‑1 lipide dans les différents tissus des bélugas (Tableau 4). Des concentrations plus faibles ont été rapportées dans les échantillons de gras provenant de biopsies (HOBBS et al., 2003) et dans les tissus d’un nouveau-né (GAUTHIER et al., 1998). McKINNEY et al. (2003) ont rapporté dans les foies des bélugas des niveaux moyens au moins dix fois plus faibles que ceux rapportés par METCALFE et al. (1998) et DEVIGNE (2003). Par ailleurs, les concentrations de Mirex chez les mâles ne semblent pas différer de celles des femelles. Enfin, aucune tendance temporelle significative n’a été rapportée pour le Mirex dans les bélugas de l’ESL (MUIR et al., 1996b, LEBEUF et al., 2007a).
L’acronyme TCPM représente les composés tris (4-chlorophényle) méthane (TCPMe) et tris (4-chlorophényle) méthanol (TCPMOH) (Figure 4E). Ces composés sont d’origine incertaine mais le TCPMOH pourrait bien être un métabolite du TCPMe. Il a été proposé que le TCPM soit un sous-produit des mélanges techniques du DDT et/ou du pesticide Dicofol (de BOER, 2000).
Le TCPM n’a été rapporté que dans le gras des bélugas (Tableau 4). Les niveaux rapportés par MUIR et al. (1996a, b) sont nettement plus élevés que ceux rapportés dans les autres études. Ces différences ont été discutées dans LEBEUF et al. (2001) et pourraient s’expliquer par une surestimation des niveaux du composé TCPMe par MUIR et al. (1996a,b) menant à une proportion de TCPMe plus élevée que celle du TCPMOH. En général, le TCPMOH est le composé dominant chez les mammifères marins, particulièrement chez les mâles (LEBEUF et al., 2001).
Une tendance temporelle du TCPMe chez le béluga de l’ESL a été rapportée par MUIR et al. (1996b) pour la période 1986-1994. Les niveaux de ce composé auraient diminué chez les femelles au cours de cette période mais pas chez les mâles. LEBEUF et al. (2007a), quant à eux, ne rapportent aucune diminution significative des niveaux de TCPM, ou encore de ses constituants, chez le béluga de l’ESL entre 1987 et 2002.
2.1.5 Les PCDD/F, non-ortho et mono-ortho BPC
Les polychlorodibenzo-p-dioxines et -dibenzofurannes (PCDD/F) sont des composés organochlorés extrêmement toxiques qui n’ont jamais été produits intentionnellement à des fins commerciales. Ces composés sont essentiellement des sous-produits générés lors de la fabrication de divers composés chlorés, notamment les BPC et le pentachlorophénol. Ils sont également produits lors de procédés thermiques et de combustion. Les plus importantes sources de PCDD/F libérés dans l’environnement sont liées à l’activité industrielle et urbaine, via les incinérateurs de déchets et les rejets d’usines chimiques et de papetières utilisant le procédé de blanchiment au chlore. Les PCDD/F possèdent une configuration planaire, ce qui constitue un facteur déterminant de leur toxicité. De plus, les congénères ayant des atomes de chlore aux positions 2, 3, 7 et 8 sont reconnus comme étant les plus toxiques (Figure 5). On compte sept composés PCDD et dix PCDF ainsi substitués, lesquels constituent les 17 congénères généralement ciblés lors des analyses chimiques. Le congénère 2, 3, 7, 8-tétrachlorodibenzo-p-dioxine (2, 3, 7, 8-TCDD) est le plus toxique.
Figure 5
Structures chimiques générales des a) polychlorodibenzo-p-dioxine (PCDD) et b) polychlorodibenzofuranne (PCDF).
General chemical structures of a) polychlorinated dibenzo-p-dioxin (PCDD) and b) polychlorinated dibenzofuran (PCDF).
(a)

(b)

Un système de calcul permet de rapporter les concentrations des différents composés PCDD/F en équivalent toxique (TEQ) de la 2, 3, 7, 8-TCDD. Cette approche a été élaborée afin de cumuler les concentrations toxiques des différents congénères de PCDD/F. Pour ce faire, la concentration de chaque congénère de PCDD/F est multipliée par son facteur d’équivalent toxique (TEF) (Van Den BERG et al., 2006). Puisque la toxicité des BPC est basée sur le même mode d’action que celle des PCDD/F, des TEF ont aussi été déterminés pour plusieurs congénères de BPC. Les BPC les plus toxiques sont ceux pouvant prendre une configuration planaire, ils n’ont pas d’atome de chlore en position ortho (non-ortho) ou un seul atome de chlore (mono-ortho) à l’une des quatre positions ortho sur la structure chimique des BPC, lesquelles réfèrent à un lien avec les atomes de carbone 2, 2’, 6 ou 6’ (Figure 1).
La présence de PCDD/F et de BPC non-ortho et mono-ortho a été déterminée dans un nombre très restreint de bélugas de l’ESL (Tableau 5). NORSTROM et al. (1992) ont été les premiers à rapporter des niveaux de PCDD/F dans dix échantillons de bélugas de l’ESL. Selon toute vraisemblance, MUIR et al. (1996b) ont rapporté, mais avec plus de détails, les niveaux de PCDD/F chez les mêmes bélugas, soit chez cinq mâles et cinq femelles récoltés entre 1987 et 1990. Les concentrations moyennes de 2, 3, 7, 8-TCDD/F étaient de l’ordre de quelques picogrammes (pg)•g‑1 lipide. Outre la 2, 3, 7, 8-TCDF, seuls quelques congénères de PCDF, lesquels n’avaient pas la substitution 2, 3, 7, 8, ont été détectés chez ces animaux alors que les niveaux de PCDD étaient systématiquement sous les limites de détection (< 1-2 pg•g‑1). Les niveaux de BPC non-ortho et mono-ortho chez les mêmes animaux étaient respectivement 103 et 106 fois plus élevés que ceux des PCDD/F. Les concentrations TEQ des PCDD/F dans le gras des bélugas de l’ESL sont négligeables par rapport à celles des BPC. Dans leur article, GAUTHIER et al. (1998) ont rapporté des niveaux de PCCD variant entre 12 et 1 170 pg•g‑1 lipide chez le béluga nouveau-né qu’ils ont examiné. Les niveaux de PCDD rapportés résultaient de la présence de deux composés fortement chlorés, un hepta et le octa-CDD alors qu’aucune PCDF n’a été détectée dans les différents tissus (gras, foie, rein et cerveau) de l’animal. Ces résultats de PCDD/F contrastent avec ceux de MUIR et al. (1996b) puisque, dans cette dernière étude, seuls des PCDF avaient été détectées. De plus, GAUTHIER et al. (1998) rapportent des niveaux de PCDD dans le gras de femelles adultes, vraisemblablement celles analysées dans les travaux de MUIR et al. (1996b). À l’instar de MUIR et al. (1996b), GAUTHIER et al. (1998) montrent que la contribution en concentrations TEQ des BPC est nettement supérieure à celle des PCDD/F, par au moins deux ordres de grandeur, dans le gras et les autres tissus examinés.
Tableau 5
Concentrations des polychlorodibenzo-p-dioxines et furannes (PCDD/F), non-ortho et mono-ortho BPC (moyenne ± écart-type, gamme) dans les tissus des bélugas du Saint-Laurent.
Concentrations of polychlorinated dibenzo-p-dioxins and furans (PCDD/F), non-ortho and mono-ortho PCB (mean ± std dev, range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (gamme) |
|
|||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
années |
TCDD/F (pg•g‑1 lip) |
PCDD (pg•g‑1 lip) |
TEQ (PCDD/F) (pg•g‑1 lip) |
Non-ortho PCB (ng•g‑1 lip) |
Mono-ortho PCB (µg•g‑1 lip) |
TEQ (PCB) (pg•g‑1 lip) |
Référence |
Gras |
C |
M5 |
37 (<1-62) |
1988-1990 |
1,4±0,2 (1,1-2,0) |
|
0,14±0,02 (0,11-0,17) |
13,1±9,0 (6,1-27,1) |
7,2±6,6 (1,8-18,2) |
1660±1300 (748-3840) |
MUIR et al., 1996a |
|
C |
F5 |
37 (5-62) |
1987-1990 |
3,0±1,0 (2,1-4,5) |
|
0,30±0,10 (0,21-0,45) |
3,2±2,4 (1,7-7,2) |
1,6±0,67 (0,62-2,2) |
380±220 (135-724) |
idem |
|
C |
?1 |
<1 |
1991 |
|
18 |
0,018 |
0,51 |
1,73 |
255 |
GAUTHIER et al., 1998 |
Foie |
C |
?1 |
<1 |
1991 |
|
1170 |
1,43 |
|
1,0 |
127 |
GAUTHIER et al., 1998 |
Rein |
C |
?1 |
<1 |
1991 |
|
261 |
0,54 |
0,17 |
0,34 |
52 |
GAUTHIER et al., 1998 |
Cerveau |
C |
?1 |
<1 |
1991 |
|
12 |
0,012 |
0,01 |
0,18 |
24 |
GAUTHIER et al., 1998 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
somme des 2,3,7,8-TCDD et 2,3,7,8-TCDF,
somme des 7 PCDD,
concentration en équivalent toxique de la 2,3,7,8-TCDD calculée en multipliant la concentration des congénères détectés par leur facteur d’équivalent toxique respectif (TEF) déterminé par NATO (1988) pour les PCDD/F et AHLBORG et al.(1994) pour les BPC,
somme des congénères non-ortho 77, 126 et 169,
somme des congénères mono-ortho 195, 118 et 156.
2.1.6 Les autres composés organochlorés
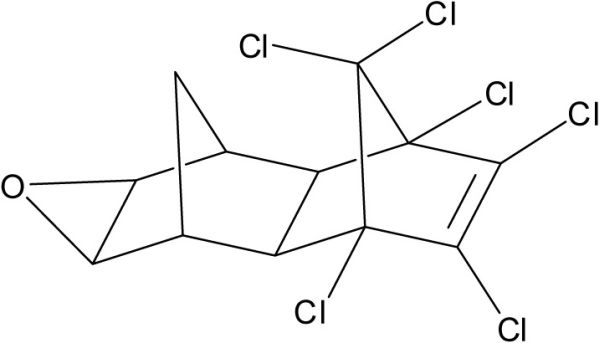
D’autres composés organochlorés ont été analysés dans les tissus des bélugas de l’ESL. Parmi ces composés, on note les polychloro-n-alkanes, la dieldrine, l’endrine, l’octachlorostyrène et certains métabolites de BPC et du 4, 4’DDE.
Les polychloro-n-alkanes (PCA), aussi nommés paraffines chlorées, sont des composés utilisés dans de nombreuses applications industrielles, notamment comme composé ignifugeant, agent plastifiant et lubrifiant pour l’industrie d’usinage métallique. Trois formulations commerciales sont courantes, selon le nombre d’atomes de carbone de la chaîne alcane, courte (C10-C13), moyenne (C14-C17) et longue (C20-C30), laquelle contient entre 30 et 70 % (poids) d’atomes de chlore (Figure 6A). Peu d’information existe sur les volumes de production des PCA et aucune réglementation n’est en place aux États-Unis et au Canada. La formulation de PCA à courte chaîne est considérée toxique au Canada (LCPE, 1993).
Figure 6
Structures chimiques du a) polychlo-n-alkane (PCA) (C10), b) dieldrine, c) endrine et d) octachlorostyrène.
Chemical structures of a) polychloro- n-alkanes (PCA) (C10), b) dieldrin, c) endrin and d) octachlorostyrene.
(a)

(b)
(c)

(d)

Deux études rapportent des niveaux de PCA dans les tissus du béluga de l’estuaire du Saint-Laurent (Tableau 6). BENNIE et al. (2000) ont rapporté des niveaux moyens de PCA, correspondant à la somme des courtes et moyennes chaînes, de 73 et 59 µg•g‑1 lipide respectivement dans le gras et le foie des carcasses de bélugas échouées entre 1986 et 1991. Ces auteurs concluent que les mâles sont généralement plus contaminés que les femelles. TOMY et al. (2000) ont rapporté des niveaux de PCA à courtes chaînes dans le gras de cinq bélugas de l’ESL récoltés au cours de la même période et dont certains animaux étaient les mêmes que ceux analysés par BENNIE et al. (2000). Les concentrations de PCA courtes chaînes rapportées par TOMY et al. (2000) étaient systématiquement de 30 à 50 fois plus faibles que celles rapportées par BENNIE et al. (2000). Selon BENNIE et al. (2000), les différences observées s’expliqueraient principalement par la méthode analytique moins sélective employée dans leur étude.
Tableau 6
Concentrations des polychloro-n-alkanes (PCA), dieldrine, endrine et octachlorostyrène (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas de l’estuaire du Saint-Laurent.
Concentrations of polychlorinated‑n-alkanes (PCA), dieldrin, endrin et octachlorostyrene (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (moyenne géométrique et/ou gamme) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
PCA (Courte C) (µg•g‑1 lip) |
PCA (Moyenne C) (µg•g‑1 lip) |
Dieldrine (µg•g‑1 lip) |
Endrine (µg•g‑1 lip) |
Octachlorostyrène (ng•g‑1 lip) |
Référence |
Gras |
C |
M4 |
35 (8-47) |
1986-1987 |
|
|
0,93±0,12 (0,81-1,1) |
|
|
MUIR et al., 1990 |
|
C |
F5 |
31 (5-58) |
1986-1987 |
|
|
(0,56±0,31) (0,21-0,87) |
|
|
Idem |
|
C |
?3 |
nd |
1952- 1986 |
|
|
0,50±0,70 (0,05-1,3) |
|
|
JARMAN et al., 1992 |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
|
|
(1,7) (0,42-5,3) |
|
(36) (<1-186) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987-1990 |
|
|
(0,63) (0,23-2,9) |
|
(63) (16-179) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
|
|
(2,0) (1,4-2,6) |
|
(53) (32-135) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
|
|
(1,4) (0,33-3,3) |
|
(39) (29-50) |
Idem |
|
C |
M10 |
35 (7-45) |
1986-1991 |
55±24 (32-100) |
47±26 (22-93) |
|
|
|
BENNIE et al., 2000 |
|
C |
F15 |
41 (3-62) |
1987-1991 |
30±22 (5-71) |
24±26 (2-93) |
|
|
|
Idem |
|
C |
M3 |
45 (40-54) |
1988-1989 |
1,0±0,55 (0,49-1,6) |
|
|
|
|
TOMY et al., 2000 |
|
C |
F2 |
54 (46-62) |
1988 |
0,84 (0,69-1,0) |
|
|
|
|
Idem |
|
B |
M34 |
14* (8-22) |
1994-1998 |
|
|
(0,60) (0,04-8,5) |
(0,26) (0,04-2,0) |
(48) (<1-508) |
HOBBS et al., 2003 |
|
B |
F10 |
14* (7-22) |
1994-1998 |
|
|
(0,20) (0,02-1,1) |
(0,16) (<0,01-0,86) |
(24) (<1-623) |
Idem |
Foie |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
|
1,1 |
0,05 |
|
METCALFE et al., 1999 |
|
C |
M3 |
27 (4-40) |
1988 |
22±8,7 (12-29) |
14±2,8 (11-17) |
|
|
|
BENNIE et al., 2000 |
|
C |
F3 |
11 (<1-32) |
1988 |
52±66 (4,9-128) |
30±35 (5,0-70) |
|
|
|
Idem |
|
C |
M3/ F3 |
40 (11-55)/ 5.3 (<1-9) |
2000-2003 |
|
|
0,76±0,19 (0,03-1,2) |
|
11±4 (<0,5-23) |
McKINNEY et al., 2006 |
Cerveau |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
|
0,55 |
0,01 |
|
METCALFE et al., 1998 |
Muscle |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
|
2,0 |
0,01 |
|
METCALFE et al., 1998 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal.
La dieldrine et l’endrine, tout comme l’aldrine, sont des insecticides chlorés non systémiques (Figure 6 B, C). Ces composés ont été utilisés au Canada pendant plus de 25 ans pour détruire les insectes dans les cultures, dans les foyers, en foresterie et dans l’industrie. L’aldrine est transformée en dieldrine par époxydation qui est elle-même plus stable et très persistante dans l’environnement. Les utilisations homologuées de ces composés ont été très limitées après le milieu des années 1970. Aux États-Unis, l’usage en agriculture de la dieldrine a été banni en 1974 et celui de l’endrine restreint en 1987. On ne signale aucune vente de ces composés au Canada depuis 1984, et à partir de 1990, leurs usages ont cessé d’être homologués.
Les concentrations moyennes de la dieldrine et de l’endrine dans les tissus des bélugas sont respectivement inférieures à 2,0 et à 0,26 µg•g‑1 lipide (Tableau 6). Comme c’est le cas pour plusieurs composés organochlorés persistants, les niveaux de dieldrine et d’endrine sont plus faibles chez les femelles. MUIR et al. (1996b) ont rapporté des concentrations stables chez les bélugas mâles entre 1986 et 1994 alors qu’elles ont diminué chez les femelles.
L’octachlorostyrène (OCS) n’est pas un composé produit à des fins commerciales mais il est plutôt généré lors de la production électrolytique du magnésium et du chlore (Figure 6D). Avant 1970, cette source de contamination de l’environnement était la plus importante alors qu’actuellement l’OCS proviendrait principalement des émissions des incinérateurs.
Les concentrations de l’OCS dans le gras et le foie des bélugas de l’ESL varient entre 10 et 60 ng•g‑1 lipide (Tableau 6). Un métabolite probable de l’OCS, le 4-OH heptachlorostyrène, a aussi été détecté dans la majorité des échantillons de foie analysés par McKINNEY et al. (2006) à des niveaux moyens de 1,5 ± 0,4 (erreur-type) ng•g‑1 lipide.
Il existe également d’autres composés organochlorés qui ont été rapportés dans les bélugas de l’ESL, notamment certains métabolites hydroxylé (OH-), et méthylsulfonylé (MeSO2‑) de congénères de BPC et du 4, 4’DDE. BERGMAN et al. (1994) ont rapporté neuf MeSO2-BPC dans le foie et le gras provenant d’une carcasse de femelle béluga récoltée sur les rives de l’ESL en 1988. Les niveaux de la ΣMeSO2‑BPC étaient respectivement de 0,4 et 0,6 µg•g‑1 lipide dans le gras et le foie, ce qui représente environ 2 % de la ΣBPC, alors qu’aucun métabolite MeSO2-DDE n’a été détecté (< 0,01 µg•g‑‑1 lipide). LETCHER et al. (2000b) ont rapporté des niveaux moyens de la ΣMeSO2‑BPC respectivement de 0,23 (gamme 0,02‑1,0) et 0,18 (0,04-0,40) µg•g‑1 lipide dans le gras récolté par la technique de biopsie chez 30 mâles et 3 femelles. Le ratio ΣMeSO2-BPC/ΣBPC était de 0,05 chez les mâles et de 0,01 chez les femelles. Ces auteurs ont également mesuré de faibles niveaux de la ΣMeSO2-DDE, notamment chez les mâles. Des niveaux nettement plus élevés de la ΣMeSO2‑BPC, en moyenne de 3,8 (± 1,3 erreur-type) µg•g‑1 lipide, ont récemment été rapportés dans le foie des bélugas de l’ESL par McKINNEY et al. (2006). Cette étude a également indiqué la présence d’un congénère MeSO2-4, 4’DDE, à des niveaux de 0,08 (± 0,04 erreur-type) µg•g‑1 lipide. Enfin, plus d’une dizaine de congénères OH-BPC ont été mesurés dans le foie des bélugas de l’ESL à des niveaux moyens de la ΣOH-BPC de 0,07 (± 0,02 erreur-type) µg•g‑1 lipide, donc très faibles par rapport à la ΣBPC (Tableau 1).
2.2 Les composés organobromés
En raison de leur caractère ignifuge, plusieurs composés organobromés sont ajoutés à divers matériaux d’usage courant. En effet, les atomes de brome permettent d’absorber l’énergie des radicaux libres qui sont générés par les flammes et ainsi retardent leur propagation. Bien que l’atome de chlore possède également cette propriété, l’atome de brome serait beaucoup plus efficace à cette fin. Malgré leur forte utilisation dans les produits commerciaux, on possède peu d’information sur les composés organobromés dans le béluga de l’ESL à l’exception d’une famille, celle des polybromodiphényles éthers (PBDE).
2.2.1 Les PBDEs et ses composés analogues
Les PBDE sont utilisés au Canada et aux États-Unis depuis de nombreuses années, cependant, les quantités utilisées jusqu’à récemment étaient relativement faibles. Depuis les années 1980, la demande à des fins commerciales pour ces composés a augmenté exponentiellement en Amérique du Nord.
Les PBDE possèdent deux cycles aromatiques pouvant contenir jusqu’à cinq atomes de brome chacun et ainsi produire 209 congénères distincts (Figure 7). On retrouve trois mélanges techniques, ou commerciaux, sur le marché : les mélanges penta, octa et déca-BDE. Le mélange penta-BDE contient majoritairement deux congénères, les BDE-47 et BDE-99 qui possèdent respectivement quatre ou cinq atomes de brome. Le mélange octa-BDE est composé principalement de deux congénères, les BDE-153 et BDE-183, qui possèdent respectivement six et sept atomes de brome, alors que le mélange déca-BDE contient presque exclusivement le congénère BDE-209, ou décabromodiphényles éther. Ces mélanges sont principalement ajoutés à divers textiles et plastiques polymériques incluant le polyuréthane, le polystyrène et l’acrylonitrile butadiène styrène (ABS). Par conséquent, ils entrent dans la fabrication de nombreux produits très courants comme les matériaux de construction et les pièces automobiles, les sous-tapis, la mousse d’ameublement et l’équipement électronique.
Figure 7
Structure chimique générale des polybro-modiphényles éthers (PBDE).
General chemical structure of polybrominated diphenylethers (PBDE).

Depuis 2004, l’unique fabricant de PBDE en Amérique du Nord, la Great Lakes Chemical Company (États-Unis), a volontairement mis un terme à la production des mélanges penta-BDE- et octa-BDE, mais continue de produire le déca-BDE. La gestion de l’usage des PBDE aux États-Unis n’est pas uniforme. La plupart des états prévoient restreindre les nouveaux usages des penta et octa-BDE ainsi que la présence de ces composés dans les matériaux à < 0,1 % en plus de suivre les tendances du déca-BDE dans l’environnement. Depuis 2003, certains états, notamment la Californie, Washington et le Maine, ont été beaucoup plus stricts à l’égard des PBDE en adoptant des réglementations visant à restreindre ou éliminer leurs usages au cours des prochaines années. Le Canada vise une élimination progressive des PBDE. En décembre 2006, les substances entrant dans la composition des trois mélanges de PBDE ont été déclarées toxiques en vertu de la Loi canadienne sur la protection de l’environnement (LCPE). La stratégie de gestion proposée par le Canada vise tous les PBDE utilisés commercialement et suggère d’interdire l’utilisation, la fabrication, la vente, la mise en vente de même que l’importation de substances qui entrent dans la composition des mélanges de penta-BDE et d’octa-BDE en plus d’interdire toute fabrication future de déca-BDE et de ses ingrédients, et de limiter la libération de déca-BDE et de ses ingrédients dans l’environnement au cours du processus de fabrication de textiles et de plastiques. Bien que le Canada ait suggéré de réglementer le déca-BDE, il ne propose pas une interdiction complète de ce type de PBDE, comme c’est le cas pour le penta-BDE et l’octa-BDE. Certaines inquiétudes ont été rapportées à l’égard d’une réglementation insuffisante dans l’usage du mélange technique déca-BDE au Canada (Ross et al., 2009).
Le tableau 7 rapporte les niveaux des principaux congénères de PBDE mesurés dans le gras et le foie des bélugas de l’ESL. On note une prédominance du congénère BDE-47 représentant à lui seul plus de la moitié des concentrations moyennes de la ΣPBDE dans le gras des bélugas examinés. Les concentrations moyennes les plus faibles ont été rapportées par LEBEUF et al. (2004) pour des animaux récoltés entre 1988 et 1999. Compte tenu de l’utilisation croissante des PBDE depuis les années 1980, il est compréhensible que les valeurs moyennes rapportées par LAW et al. (2003) et McKINNEY et al. (2006) soient plus élevées puisque ces études rapportent des niveaux chez des animaux récoltés récemment. Toutefois, les niveaux de PBDE dans le foie des bélugas de l’ESL (McKINNEY et al., 2006) semblent plus importants que ceux dans son gras (LAW et al., 2003). Selon les observations rapportées par McKINNEY et al. (2006), les niveaux de PBDE chez les jeunes animaux sont nettement plus élevés, jusqu’à un ordre de grandeur, par rapport à ceux mesurés chez les adultes. Ceci suggère que les concentrations relativement élevées de PBDE rapportées par McKINNEY et al. (2006) dans le foie des bélugas de l’ESL s’expliqueraient par l’âge des animaux examinés plutôt que par le tissu analysé. Selon GEBBINK et al. (2008), l’enrichissement des PBDE dans le foie des mammifères, notamment l’ours polaire (Ursus maritimus), pourrait être en partie expliqué par l’affinité de ces composés pour certaines protéines endogènes qui s’accumulent préférentiellement dans le foie par rapport au tissu adipeux. La contamination en PBDE rapportée chez les bélugas mâles et femelles est très semblable. LEBEUF et al. (2004) explique cette situation par un taux très élevé d’augmentation des niveaux des PBDE chez les deux genres qui viendrait masquer l’élimination attendue des PBDE chez les femelles lors de la gestation et la lactation. En effet, les concentrations de PBDE ont augmenté exponentiellement chez les bélugas de l’ESL entre 1988 et 1999 en doublant à tous les 2-3 ans au cours de cette période. (LEBEUF et al. 2004).
Tableau 7
Concentrations des polybromodiphényles éthers (PBDE) (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas du Saint-Laurent.
Concentrations of polybrominated diphenylethers (PBDE) (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (gamme) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
BDE-47 (µg•g‑1 lip) |
BDE-99 (µg•g‑1 lip) |
BDE-100 (µg•g‑1 lip) |
Σ5PBDE (µg•g‑1 lip) |
ΣPBDE (µg•g‑1 lip) |
Référence |
Gras |
C |
M7 |
29 (18-38) |
1999-2000 |
0,28 |
0,11 |
0,06 |
|
0,54 |
LAW et al., 2003 |
|
C |
F8 |
30 (17-54) |
1999-2000 |
0,26 |
0,10 |
0,06 |
|
0,54 |
Idem |
|
C |
M28 |
39 (18-58) |
1988-1999 |
0,15±0,11 (0,01-0,39) |
0,05±0,05 (<0,01-0,20) |
0,03±0,03 (<0,01-0,09) |
0,28±0,20 (0,03-0,73) |
0,29±0,21 (0,03-0,78) |
LEBEUF et al., 2004 |
|
C |
F26 |
42 (20-63) |
1988-1999 |
0,17±0,15 (0,01-0,54) |
0,07±0,06 (<0,01-0,22) |
0,04±0,04 (<0,01-0,13) |
0,33±0,28 (0,02-1,0) |
0,34±0,29 (0,02-1,1) |
Idem |
Foie |
C |
M3/ F3 |
40 (11-55)/ 5,3 (<1-9) |
2000-2003 |
|
|
|
2,0±0,43 (0,21-2,8) |
2,20±0,66 (0,25-3,0) |
McKINNEY et al., 2006 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
congénères BDE‑47, ‑99, ‑100, ‑153 et ‑154
En plus des congénères de PBDE, lesquels proviendraient essentiellement des mélanges techniques, McKINNEY et al. (2006) ont rapporté la présence de composés analogues aux PBDE chez le béluga de l’ESL. Ces composés possèdent un groupe fonctionnel méthoxyle (MeO-) et sont mesurés à des niveaux (ΣMeO-PBDE) variant entre 20 et 25 ng•g‑1 lipide dans le foie des bélugas. Parmi les 15 composés MeO-PBDE recherchés, seulement deux composés, dont le 6-MeO-BDE-47, ont été détectés (McKINNEY et al., 2006). LEBEUF et al. (2005) ont également rapporté la présence de MeO-PBDE dans le biote de l’ESL, notamment chez le poulamon Atlantique (Microgadus tomcod) a des concentrations hépatiques environ deux fois supérieures à celles mesurées dans le foie des bélugas. Dans le Saint-Laurent, l’origine des MeO-PBDE n’est pas connue. Cependant, il a été démontré qu’aux moins deux MeO-PBDE, incluant le 6-MeO-BDE-47, accumulés par une baleine à bec de True (Mesoplondon merus) de l’Atlantique du Nord, seraient d’origine naturelle, et donc ne proviendraient pas de la transformation des PBDE d’origine anthropique (TEUTEN et al., 2005).
3. Interprétation des niveaux et des tendances des polluants organiques peristants chez le béluga du Saint-Laurent
À la suite de premiers travaux de SERGEANT (1980), une vingtaine d’études ont rapporté des niveaux de polluants organiques persistants dans divers tissus des bélugas du Saint-Laurent. Cependant, l’interprétation des résultats rapportés dans ces études, et surtout la comparaison des résultats entre les études elles-mêmes, comportent certains défis. En effet, plusieurs facteurs et conditions influencent directement, et souvent d’une manière importante, les concentrations et les tendances temporelles des contaminants accumulés chez le béluga de l’ESL. Les mêmes difficultés s’appliquent également lors de la comparaison des concentrations et des tendances temporelles de la contamination du béluga de l’ESL avec celles d’autres populations de bélugas ou d’autres espèces de cétacés.
3.1 Méthodes utilisées et manière de rapporter les résultats d’analyses
Au cours des ans, les méthodes analytiques utilisées pour doser les POP se sont grandement améliorées, notamment en ce qui concerne la séparation des composés par chromatographie gazeuse en passant de la colonne garnie à la colonne capillaire au cours des années 1980. De plus, l’identification et la confirmation des pics chromatographiques aux composés d’intérêt sont devenus beaucoup plus fiables à l’aide de détecteur de masse de type quadripôle ou trappe ionique. Enfin, la disponibilité sur le marché des composés d’intérêt authentiques et de matériaux de référence a contribué à améliorer et valider les méthodes d’analyses. Une des plus importantes conséquences de ces améliorations a été le passage de la quantification de familles de composés en référence au patron chromatographique des composés présents dans les mélanges techniques, notamment pour les BPC, toxaphène et PCA, à celle des composés ou congénères individuels.
Malgré ces avancements technologiques, il est encore aujourd’hui très commun de rapporter les concentrations de la somme des composés constituant une famille de POP, comme c’est généralement le cas pour la ΣDDT, les HCH et CHL, plutôt que celles des composés individuels. Considérant que le nombre de composés analysés dans chaque famille de POP peut varier d’un laboratoire à un autre, et qu’il augmente généralement aux cours des ans, la comparaison des niveaux entre les études demeure difficile. Le défi est encore plus important lorsque les résultats de plusieurs études sont rassemblés afin de déterminer les tendances temporelles. Enfin, la façon de rapporter les résultats d’analyses, soit à l’aide de moyennes arithmétiques ou géométriques, ou encore sur une base de poids humide ou de poids lipidique, fait en sorte que la comparaison des résultats de polluants organiques persistants rapportés chez le béluga de l’ESL n’est pas simple.
3.2 Les caractéristiques des bélugas, les tissus analysés et leur qualité
Plusieurs caractéristiques des bélugas sont susceptibles d’influencer les niveaux et les tendances des polluants organiques persistants chez les individus de la population de l’ESL. La première caractéristique à considérer est sans doute le sexe des animaux et son impact sur la charge accumulée de polluants organiques persistants. En effet, les femelles sont généralement moins contaminées que les mâles en raison de leur capacité à transférer certains POP à leur petit lors de la gestation et de la longue période de lactation qui dure près de deux ans. En rapportant une contamination significative chez un béluga nouveau-né, GAUTHIER et al. (1998) supporte l’importance du transfert gestationnel des POP. Selon Hickie et al. (2000), la lactation constitue une voie importante de transfert des POP qui mène les veaux à atteindre des sommets de contamination. Il faut cependant noter que certains composés sont peu transférés, notamment le Mirex qui a été mesuré à de très faibles niveaux chez le béluga nouveau-né, ce qui expliquerait les concentrations similaires chez les mâles et les femelles (Tableau 4). On note également un ratio 4, 4’DDE/ΣDDT généralement plus faible chez les femelles que chez les mâles (Tableau 2). Ceci pourrait possiblement s’expliquer par un transfert préférentiel du 4, 4’DDE vers le petit par rapport aux autres constituants. On observe, en effet, que chez le nouveau-né (Gauthier et al., 1998) la somme des DDT est constituée à près de 90 % de 4, 4’DDE. En plus de cette capacité à transférer des contaminants à son veau, la femelle béluga se distingue du mâle notamment dans la fréquentation d’habitat estival distinct (MICHAUD et al., 1993). Cette différence de fréquentation d’habitat expliquerait possiblement les ratios d’isotopes stables distincts entre les mâles et les femelles rapportés par LESAGE et al. (2001). Selon LEBEUF et al. (2007a), les différences de niveaux de POP entre les mâles et les femelles s’expliquent principalement par le transfert de contaminants vers le veau lors de la gestation et de la lactation. Cependant, les différences observées dans les tendances temporelles entre les mâles et les femelles doivent faire intervenir d’autres facteurs, notamment la fréquentation d’habitat distinct conduisant vraisemblablement à une diète différente.
La longévité du béluga est également une caractéristique de l’espèce qui influence sa charge de contaminants. Selon HICKIE et al. (2000), le béluga mâle verrait sa contamination augmenter continuellement à partir de la fin de sa croissance physique, alors que chez les femelles, la contamination atteindrait un plateau durant la période de reproduction pour augmenter de nouveau vers la fin de sa vie. Bien que les changements de contamination au cours de la vie des bélugas soient généralement acceptés, de telles relations ne sont pas toujours clairement démontrées. En effet, MARTINEAU et al. (1987) rapportent des augmentations de BPC et de la ΣDDT en fonction de l’âge chez les mâles et les femelles alors que MUIR et al. (1996a) montrent des corrélations négatives des BPC et CHL avec l’âge chez les mâles et des relations positives chez les femelles. Une des grandes faiblesses dans l’établissement d’une relation entre la contamination et l’âge des animaux découle du manque de connaissances de l’intensité de l’exposition des bélugas aux POP au cours de la vie des animaux, incluant lors du transfert maternel. En effet, les animaux qui ont vécu entre les années 1960 et 1980 ont fait face à des changements importants dans leur exposition aux contaminants organiques persistants, notamment lors du transfert maternel de contaminants. Ainsi, il semble assez difficile d’évaluer les effets de l’âge sur la contamination des animaux de longue durée de vie sans corriger pour les changements temporels de leur exposition. On peut penser que la forte contamination par les PBDE chez les jeunes bélugas par rapport aux adultes (McKINNEY et al., 2006) est une indication de l’augmentation récente de l’exposition des bélugas à ces composés reflétée par un transfert maternel actuel plus important vers les jeunes qu’il ne l’a été vers ses parents.
Dans leurs travaux, HOBBS et al. (2003) ont comparé les concentrations de POP dans des échantillons de gras prélevés sur des bélugas vivants, à l’aide de la technique de biopsie, à celles mesurées dans des échantillons prélevés sur des carcasses. Ce travail visait à évaluer si un changement de conditions des animaux au terme de leur vie, telle l’émaciation, a une influence majeure sur le niveau de leur contamination. Afin de réduire les biais entre les deux groupes, uniquement les mâles ont été considérés et les animaux sélectionnés avaient un âge moyen très semblable. Cependant, depuis cette étude, la méthode de détermination de l’âge des bélugas à partir de la dentine, appliquée spécifiquement aux animaux échoués, a été revue et corrigée de manière à ce que les carcasses de bélugas considérées étaient en réalité deux fois plus âgées que les animaux biopsiés. Il est clair que le biais potentiel créé par une telle différence d’âge entre les groupes d’animaux rend les résultats de la comparaison de la contamination entre les animaux vivants et les carcasses questionnables. Il n’existe actuellement aucune étude portant sur le béluga de l’ESL qui vienne démontrer l’influence de la condition des animaux sur leurs niveaux de contamination. Cependant, d’autres études ont tenté de répondre à cette question en comparant la contamination de mammifères marins dont la cause de la mort variait entre deux extrêmes, la mort accidentelle ou la mort naturelle. Selon les travaux de KUIKEN et al. (1994), les concentrations de POP mesurées dans les carcasses de marsouins communs (Phocoena phocoena) échouées sur les côtes britanniques n’étaient pas différentes de celles des animaux morts accidentellement par noyade lors d’activité de pêche. Cependant, d’autres études montrent que la contamination des carcasses échouées, particulièrement celles dont la cause de la mort est une maladie infectieuse, est généralement plus élevée que celle des animaux morts accidentellement (JEPSON et al., 1999, 2005).
Les contaminants organiques persistants lipophiles s’accumulent préférentiellement dans le gras et les tissus riches en lipide. Une façon courante de comparer les concentrations de POP entre différents tissus consiste à rapporter les niveaux sur une base de poids lipidique du tissu analysé. Cependant, une application inadéquate de cette approche peut induire certains biais (HEBERT et KEENLEYSIDE, 1995). En général, les études ont rapporté des concentrations de contaminants chez les bélugas de l’ESL dans un seul tissu, le gras. Occasionnellement, le foie, le rein, le poumon, le lait, le cerveau ainsi que le muscle ont été analysés chez certains animaux. Seuls MARTINEAU et al. (1987) ont comparé la contamination entre différents tissus, notamment le gras, le foie et le rein, chez un nombre substantiel (n = 17) de bélugas. Ces auteurs concluent que les concentrations de POP, rapportées sur une base de poids lipidique, sont assez semblables entre les tissus bien que des cas extrêmes soient observés. Par ailleurs, les études portant sur les tendances temporelles de la contamination du béluga de l’ESL ont à ce jour utilisé exclusivement le gras, un tissu qui pourrait ne pas très bien refléter la contamination des tissus internes des animaux. Il serait pertinent de comparer les tendances temporelles des POP chez le béluga de l’ESL dans un tissu interne, notamment le foie, et de comparer ces tendances avec celles rapportées dans le gras.
La représentativité du tissu analysé constitue une préoccupation importante lorsque vient le temps de comparer les résultats rapportés par des études utilisant des techniques d’échantillonnage différentes. Par exemple, le gras provenant d’une biopsie représente la couche superficielle du gras, alors que le prélèvement sur une carcasse permet d’intégrer le gras sur toute son épaisseur, laquelle peut atteindre 10 cm. LEBEUF et al. (2000) ont sectionné le gras de quatre bélugas de l’ESL en sous-couches de gras de 1 cm d’épaisseur et analysé les BPC dans chaque sous-couche afin d’examiner si il y a stratification de la contamination. Ils ont conclu que la distribution de plusieurs congénères de BPC était homogène sur toute la profondeur du gras alors que pour d’autres congénères, des gradients, quelques fois inverses, ont été observés. Plus récemment, LEBEUF et al. (2005) rapportaient que la gamme de concentrations de POP observée dans les sous-couches de gras chez un béluga était du même ordre de grandeur que les variations de concentrations observées entre différents sites de prélèvement sur le même animal. La littérature scientifique fournit peu d’information sur la stratification des POP dans le gras des cétacés à dents (odontocètes).
Dans le cas spécifique de prélèvements de tissus sur des carcasses, la qualité de la carcasse, et par conséquent celle des tissus récoltés, peut vraisemblablement créer un biais dans les concentrations mesurées. En pratique, la qualité des tissus n’est généralement pas considérée lors d’exercice de comparaison de la contamination par les POP entre groupes d’animaux puisqu’il n’existe pas suffisamment d’études pour tirer de solides conclusions sur ce biais potentiel. Dans le cadre du programme des carcasses de bélugas de l’ESL, chaque carcasse est codée en appliquant l’échelle de GERACI et LOUNSBURY (2005). Une évaluation préliminaire a permis de démontrer qu’il n’y avait pas de corrélation entre la contamination par les POP mesurée dans le gras du béluga et le code de qualité de sa carcasse (LEBEUF, données non publiées). Ces résultats sont supportés par les faibles variations (moyenne ± écart-type; médiane) du contenu lipidique dans les échantillons de gras chez les mâles (85,9 ± 7,0 %; 91,3 %; n = 39) et les femelles (96,6 ± 4,7 %; 94,1 %; n = 38) rapportées dans l’article de LEBEUF et al. (2007a).
Dans le but d’éviter de créer un biais lors de la comparaison des concentrations ou de la caractérisation des tendances temporelles de la contamination chez le béluga, il est souhaitable de considérer des individus ayant des caractéristiques semblables. En général, les comparaisons de niveaux se font en considérant un seul genre à la fois et en sélectionnant des groupes d’animaux ayant un âge moyen semblable. Dans le cas des études de tendances temporelles, il est recommandé de ne pas combiner les échantillons provenant d’animaux vivants avec ceux provenant de carcasses échouées. Il est également nécessaire de démontrer qu’il n’y a pas de changements temporels significatifs de différentes caractéristiques chez les animaux sélectionnés. Dans leur étude, LEBEUF et al. (2007a) ont validé qu’il n’y avait aucune tendance significative de l’âge, de la longueur, de l’épaisseur de la couche de gras et de la qualité de la carcasse chez les animaux analysés pour la période considérée dans leur évaluation des tendances temporelles de la contamination du béluga. En plus des caractéristiques énumérées précédemment, il se pourrait qu’à l’intérieur de la population des bélugas de l’ESL, certains groupes d’animaux se distinguent par leur aire de haute fréquentation (habitation et alimentation). De telles différences entre groupes d’animaux, extrêmement difficiles à déterminer à partir des carcasses, pourraient possiblement expliquer une partie de la forte variabilité dans la contamination observée entre les individus et influencer les tendances temporelles évaluées sans tenir compte de ce facteur.
3.3 Les propriétés et caractéristiques des POP
À l’intérieur même d’une famille de polluants organiques persistants, les composés possèdent certaines propriétés pouvant couvrir une gamme importante de valeurs qui différencient leur comportement spécifique. Par exemple, le transfert sélectif des PBDE lors de lactation chez le phoque à capuchon (Cystophora cristata) est très fortement corrélé avec le caractère lipophile des composés, représenté par leur coefficient Kow (LEBEUF et al., 2007b). L’accumulation des POP, qui se fait principalement via la nourriture chez les mammifères marins, est généralement considérée efficace et peu sélective alors que l’élimination des composés serait beaucoup plus sélective, une conséquence probable de la réduction de la mobilité des composés les plus lipophiles accumulés dans les organismes. En effet, LEBEUF et al. (2007a) montrent que le taux d’élimination des POP diminue avec l’augmentation du Kow des composés accumulés chez le béluga de l’ESL. Ainsi, certaines caractéristiques chimiques des POP, tel le Kow, ont une influence sur les niveaux et tendances temporelles des POP observés chez le béluga et possiblement les distinguent de ceux observés dans leur habitat. La capacité de certains POP à se lier à certaines protéines en raison de la présence d’un groupe fonctionnel ou d’une conformation structurale spécifique constitue un facteur supplémentaire qui influence leur accumulation chez les organismes.
Malgré que les POP répondent tous aux critères de persistance dans l’environnement, certains sont plus sensibles que d’autres à une transformation biochimique par les organismes du milieu. Les mammifères marins possèdent une capacité métabolique accrue par rapport à d’autres organismes, ce qui les mène, notamment, à modifier le patron de BPC acquis via leur diète tel qu’il est rapporté par LEBEUF et al. (1999) chez quatre espèces de phoques de l’ESL. Selon BERNT et al. (1999), la différence de patrons de BPC entre le béluga et le phoque commun (Phoca vitulina) résidant de l’ESL s’expliquerait par leur capacité métabolique distincte. En effet, il a été démontré que les phoques possèdent une capacité accrue à métaboliser certains congénères de BPC par rapport aux petits cétacés (BOON et al., 1997; TANABE et al., 1988). Bien que les différences de capacité métabolique entre les espèces puissent influencer le patron de POP qu’elles accumulent, on doit aussi considérer l’influence d’une différence de diète entre les espèces provenant du même habitat pour expliquer les différences de patrons de contaminants accumulés. D’une manière similaire, les différences de patrons de POP observées entre le béluga de l’ESL et ceux de l’Arctique canadien résulteraient vraisemblablement des différences de diète entre les populations. Il a cependant été suggéré que la présence de niveaux élevés de contaminants chez le béluga de l’ESL était susceptible d’induire l’activité métabolique à un degré supérieur chez cette population par rapport aux populations des bélugas de l’Arctique. LEBEUF et al. (2007a) rapporte que le béluga de l’ESL non seulement possède la capacité métabolique mais qu’il l’exerce en transformant certains BPC, notamment ceux du groupe structural III, selon la classification de BOON et al. (1994), qui regroupe les congénères possédant au plus un atome de chlore en position ortho (positions 2, 2’, 6, 6’, Figure 1) et des atomes d’hydrogène adjacents uniquement aux positions ortho et méta (positions 3, 3’, 5, 5’, Figure 1). En plus de diminuer les concentrations de certains POP par des mécanismes de biotransformation, ce type de réaction peut aussi conduire à l’enrichissement de produits de transformation dans les organismes. En effet, dans leur étude sur la transformation de PBDE chez la carpe commune (Cyprinsu carpio) STAPLETON et al. (2004) ont démontré que certains congénères de PBDE produisent à la suite d’une réaction de débromination des congénères persistants, notamment le BDE-47. En somme, les concentrations et les tendances temporelles de certains contaminants observées chez les bélugas de l’ESL sont aussi influencées par la capacité de biotransformation de l’espèce.
3.4 Changements dans l’habitat du béluga de l’ESL
Les changements dans la production et les usages des POP au Canada et aux États-Unis ont généré d’importants changements d’apports de POP dans l’habitat du béluga de l’ESL. En effet, l’usage de plusieurs POP a été réglementé, voire banni, au cours des 40 dernières années, ce qui a conduit à une diminution générale des niveaux dans les sédiments de l’ESL (LEBEUF et NUNES, 2005). D’autres composés, notamment les PBDE, ont connu une augmentation des usages qui a conduit inévitablement à une augmentation de la contamination de l’habitat du béluga, si on se fit à l’augmentation des niveaux de PBDE observés chez le béluga lui-même (LEBEUF et al., 2004). Il semble donc raisonnable de croire que les tendances de la contamination des POP observées chez le béluga de l’ESL reposent principalement sur les changements dans la production et les usages de ces composés au Canada, aux États-Unis et ailleurs sur la planète puisque les POP se déplacent sur de longues distances par voie atmosphérique et, dans une moindre mesure, océanique.
Au cours des ans, l’habitat du béluga a fait face à d’autres changements importants, notamment au niveau de sa structure biologique. En effet, plusieurs stocks de poissons, incluant l’anguille d’Amérique (Anguilla rostrata), l’éperlan arc-en-ciel (Osmerus mordax), le poulamon atlantique (Microgadus tomcod) et la morue franche (Gadus morhua) ont connu des chutes drastiques dans l’estuaire du Saint-Laurent (CASTONGUAY et al., 1994; MYERS et al., 1997). L’effondrement de certains stocks de poissons a possiblement eu un impact sur la diète du béluga de l’ESL et, par conséquent, sur sa contamination. En effet, à l’aide de leur modèle, HICKIE et al. (2000) prédisaient une diminution accélérée des niveaux de BPC chez le béluga de l’ESL si l’anguille d’Amérique était progressivement exclue de sa diète. Ainsi, on ne peut ignorer l’influence des changements survenus dans l’habitat du béluga et leurs conséquences sur les concentrations et les tendances temporelles de la contamination par les POP observées chez le béluga de l’ESL.
4. Les autres composés auxquels est exposé le béluga
En plus d’accumuler des concentrations importantes et une grande diversité de polluants organiques persistants, le béluga de l’estuaire du Saint-Laurent est exposé à d’autres composés dont les caractéristiques physicochimiques, notamment leur stabilité, et les sources diffèrent de celles des POP. Ces composés s’accumulent à divers degrés chez le béluga.
4.1 Les autres composés persistants
Plusieurs autres groupes de composés persistants, dont la structure chimique n’est pas organique, c’est-à-dire constituée principalement d’atomes de carbone et d’hydrogène, ou entièrement organique, ont été mesurés dans les tissus des bélugas de l’ESL.
Deux études ont rapporté des niveaux de tributylétain (TBT), un composé organo-métallique retrouvé dans les agents antisalissures utilisés pour peindre la coque des bateaux. YANG et al. (1998) ont mesuré des niveaux de TBT et de ses produits de transformation, le dibutylétain (DBT) et le monobutylétain (MBT), dans le foie et le gras de carcasses de bélugas récoltées en 1987 et 1988. Selon ces auteurs, les concentrations de la somme des butylétain, ΣBT, dans le foie étaient en moyenne de 560 (± 513 écart-type) ng Sn•g‑1 poids sec du tissu soit environ dix fois plus élevées que dans le gras. ST-LOUIS et al. (2000) ont rapporté des niveaux moyens semblables, de 375 (± 477 écart-type) ng Sn•g‑1 poids sec, dans le foie de carcasses de bélugas récoltées entre 1995 et 1998. Les deux études montrent que le DBT est généralement prédominant chez les bélugas sauf pour de très jeunes bélugas chez qui le TBT domine et pour quelques bélugas adultes parmi les plus âgés chez qui les concentrations de MTB sont les plus élevées.
Certains métaux, incluant le cuivre, le plomb, le cadmium, le mercure, le sélénium et le zinc, ont été analysés dans le foie, le rein et le muscle de carcasses de bélugas de l’ESL récoltées entre 1982 et 1986 (WAGEMANN et al., 1990). Les concentrations de plomb, mercure (Hg) et sélénium seraient particulièrement élevées chez les bélugas de l’ESL par comparaison aux bélugas de populations arctiques. SERGEANT(1980) a rapporté des niveaux de Hg de 123 µg•g‑1 poids sec (après conversion) dans le foie d’une femelle béluga adulte dont la carcasse a été récoltée en 1979, ce qui est semblable aux niveaux moyens de 126 (± 161 écart-type) µg•g‑1 poids sec rapportés par WAGEMANN et al. (1990) chez 30 bélugas de l’ESL.
Le fluor fait partie de la série chimique des halogènes tout comme le chlore ou le brome. Le fluor est également utilisé dans certaines applications industrielles, notamment par l’industrie de l’aluminium et des pâtes et papiers. Des niveaux élevés de fluor peuvent provoquer divers types de fluorose, dont les fluoroses dentaire et osseuse. MIKAELIAN et al. (1999) ont rapporté des concentrations moyennes de fluor de 4 500 (± 900 écart-type) µg•g‑1 poids sec dans les os des bélugas de l’ESL. Ces niveaux sont semblables à ceux rapportés chez d’autres populations de baleine et ne montrent pas de signe clinique de fluorose chez les animaux examinés. Chez les humains, les premiers signes cliniques de fluorose osseuse se manifestent à des concentrations de 6 000 µg de fluor par gramme de cendre d’os.
Plutôt que d’analyser divers composés halogénés spécifiques dans les tissus des bélugas, KICENIUK et al. (1997) ont mesuré par activation neutronique la charge en atomes d’halogène associée à la matière organique extraite de divers tissus. Ainsi, le contenu en chlore, brome et iode a été rapporté dans le cerveau, le rein et le foie de six à huit bélugas de l’ESL dont les carcasses ont été récoltées en 1988 et 1989. Les niveaux de chlore extrait, exprimés sur une base lipidique, sont variables entre les tissus, favorisant le cerveau chez la population de l’ESL et le rein chez la population de l’Arctique étudiée.
Les concentrations de quatre composés diméthyles bipyrroles halogénés, dont l’origine est vraisemblablement naturelle, ont été rapportées dans le gras des bélugas de l’ESL par TITTLEMIER et al. (2002). Les niveaux moyens de 14,8 (gamme 2,4-52,7) ng•g‑1 lipide étaient relativement faibles, soit environ 1 000 fois inférieurs à ceux du congénère de BPC le plus prédominant dans les mêmes échantillons, le BPC-153.
Plusieurs composés organiques persistants n’ont pas été analysés dans les tissus des bélugas de l’ESL malgré leur présence probable. En effet, tous les composés mesurés dans les tissus des bélugas ont aussi été rapportés dans le biote des Grands Lacs, notamment celui du lac Ontario. Sur cette base, on peut penser que le béluga de l’ESL accumule divers composés organochlorés incluant les polychlorodiphényles éthers (PCDE) et les polychloronaphtalènes (PCN) (Helmet al., 2006; NIIMI et al., 1994), divers composés organobromés incluant l’hexabromocyclodo-décane (HBCD), le 1,2-bis(2,4,6-tribromophénoxy)éthane (BTBPE), le 1,2,3,4,5-pentabromoéthylbenzène, l’hexabromobenzène (HBB), le pentabromoéthylbenzène (PBEB), le pentabromotoluène (PBT), le déchlorane plus (DP) (Gauthieret al., 2007; Hiteset al., 2006) ainsi que certains composés organofluorés incluant deux grands groupes de composés perfluorés, les acides perfluoroalkyle sulfoniques dont le perfluorooctane sulfonate (PFOS), et les acides perfluoro-carboxyliques dont l’acide perfluoro-octanoïque (PFOA) (GIESY et al., 2006; HOUDE et al., 2006). Des données préliminaires confirment la présence du PFOS dans le foie des bélugas de l’ESL à des concentrations moyennes de 10 µg•g‑1 lipide (LEBEUF, données non publiées).
4.2 Les composés non bioaccumulés
Certains composés chimiques d’origine anthropique ne s’accumulent pas, ou très peu, dans les organismes aquatiques. Cependant, ces composés peuvent être présents dans le milieu aquatique et constituer une exposition continue et perturbatrice pour les organismes.
Parmi ces composés, mentionnons les nouveaux pesticides ainsi que certains composés pharmaceutiques. Ces composés chimiques sont soit très instables dans l’environnement, et par conséquent leur action est de courte durée, ou soit que leurs caractéristiques physicochimiques ne favorisent pas leur rétention dans les organismes.
Il existe aussi des composés chimiques qui s’accumulent dans l’environnement, notamment dans les sédiments, mais qu’on ne retrouve pas dans les organismes tels que les poissons et les mammifères marins en raison de la capacité de ces derniers à les métaboliser. Les hydrocarbures aromatiques polycycliques (HAP) constituent un bon exemple de ce type de composés. En effet, il est reconnu qu’une partie de l’habitat du béluga de l’ESL a été fortement contaminée par les HAP suite aux activités industrielles locales (MARTEL et al., 1986). Bien que le béluga ait été et continue d’être exposé aux HAP, ces composés ne s’accumulent dans ses tissus. Selon MARTINEAU et al. (2002), l’exposition du béluga de l’ESL aux HAP, notamment au benzo-α-pyrène, serait liée à l’étiologie des cancers observés chez cette population.
5. Perspectives
Après plus de 20 articles scientifiques publiés sur la contamination du béluga de l’estuaire du Saint-Laurent par divers polluants organiques persistants, que retire-t-on de ces informations dans la perspective de la contamination de cette population et de celle de son habitat?
Les études réalisées sur la contamination du béluga de l’ESL permettent d’obtenir une bonne, voire excellente, vue d’ensemble de la diversité de la contamination par les composés organiques persistants chez cette population. Cependant, la majorité des études rapporte des concentrations de POP mesurées chez des bélugas récoltés sur une courte période temps, généralement quelques années. En raison des facteurs qui diffèrent entre ces études spécifiques et influencent l’accumulation de POP (voir section 3), il est difficile de tirer de solides conclusions à partir de la comparaison de la contamination des bélugas de l’ESL rapportée dans ces études ou de combiner les résultats de ces études afin d’évaluer les tendances temporelles de cette contamination. La comparaison de la contamination entre différentes populations de bélugas ou d’autres mammifères marins souffre généralement des mêmes difficultés. Il faut donc interpréter les résultats de ce type de comparaison avec précaution et n’en tirer que les grandes évidences. C’est en effet ce qu’ont fait HOUDE et al. (2005) dans leur article de synthèse en comparant les concentrations de POP chez les cétacés (Delphinoïdes). En bref, les auteurs rapportent que la population du béluga de l’ESL est généralement plus contaminée par les POP que les populations des bélugas de l’Arctique. Cependant, la contamination du béluga de l’ESL se situe à un niveau intermédiaire par rapport à celle d’autres cétacés habitant diverses parties du monde.
Afin de suivre adéquatement les changements temporels de la contamination du béluga de l’ESL, il est nécessaire de procéder à un échantillonnage régulier de tissus chez un nombre assez important d’animaux. Depuis 1997, notre équipe récolte systématiquement des tissus de bélugas pour des fins d’analyses de contaminants en collaboration avec le programme des carcasses. En moyenne, la récolte de tissus s’effectue sur 15 bélugas échoués par année (MEASURES, 2008). Certains échantillons récoltés avant 1997 et archivés ont permis d’élargir les périodes couvertes dans les études de suivi de la contamination du béluga réalisées à l’Institut Maurice-Lamontagne, Pêches et Océans Canada (GOUTEUX et al., 2003; LEBEUF et al., 2004; LEBEUF et al., 2007a). Le suivi de la contamination du béluga de l’ESL constitue donc une suite logique du programme des carcasses du béluga de l’ESL en permettant de déterminer les tendances des concentrations de polluants organiques persistants accumulées dans les tissus des individus de cette population menacée. Il faut cependant retenir que les tissus analysés proviennent de carcasses d’animaux et, par conséquent, l’extrapolation de la contamination mesurée dans les tissus des carcasses à celle de la population vivante n’est ni directe, ni évidente.
Bien qu’imparfait, le suivi de la contamination chez les bélugas de l’ESL constitue un outil d’évaluation de l’impact des mesures visant à réduire les apports de ces composés et les perturbations dans l’habitat du béluga. Malgré une diminution des niveaux de certains composés réglementés, la population du béluga de l’ESL demeure fortement contaminée par les POP. De plus, des augmentations récentes de concentrations de composés émergents, notamment les PBDE, ont été rapportées chez le béluga. En somme, la charge nette en composés toxiques accumulés dans les tissus des bélugas de l’ESL continue à être élevée alors que sa diversité augmente. Ces résultats suggèrent que la contamination des bélugas par les POP continue de présenter des risques d’effets toxiques sur la population.
Un des principaux buts de la caractérisation et du suivi temporel de la contamination du béluga de l’ESL est de tenter d’établir un lien entre les concentrations des composés toxiques dans les individus et les effets néfastes qu’elles sont susceptibles de produire. Il semble maintenant possible de tirer avantage des connaissances acquises au cours des 20 dernières années afin d’explorer l’existence d’un lien quantifiable entre la contamination du béluga par les POP et la cause probable de sa mort telle que déterminée lors de la nécropsie réalisée sur sa carcasse. En effet, chez le marsouin commun, une association a récemment été démontrée entre sa contamination par les BPC et l’incidence des mortalités causées par une maladie infectieuse (JEPSON et al., 2005). Une telle association chez le béluga de l’ESL viendrait supporter l’hypothèse que le système immunitaire de certains bélugas de l’ESL est affecté par la présence des POP et a un effet négatif sur la population (BROUSSEAU et al., 2003). La précarité de la population des bélugas de l’ESL pourrait aussi être liée à son exposition à des composés chimiques qui ne s’accumulent pas chez l’espèce, tel que l’a suggéré MARTINEAU et al. (2002). Il est donc essentiel d’acquérir plus d’informations sur l’exposition globale des bélugas à divers types de contaminants. Les futurs travaux de recherche devront permettre d’optimiser les efforts de suivi de la contamination dans les tissus du béluga, dans sa diète et dans son habitat vers les composés, ou les groupes de composés, les plus susceptibles de causer des effets nocifs sur la santé et la reproduction de cette population (BÉLAND et al., 1993). Dans cette perspective, il sera aussi nécessaire de poursuive le développement d’outils toxicologiques permettant d’identifier les effets de l’exposition des bélugas et de ses proies aux contaminants présents dans son habitat. La présence de contaminants dans l’habitat du béluga peut avoir des effets délétères chez divers organismes du milieu et ainsi affecter indirectement le béluga de l’ESL, notamment au niveau des quantités mais aussi de la qualité des proies disponibles pour son alimentation. Il devient essentiel d’opter pour une approche globale, dite écosystémique, dans l’étude du béluga et l’application d’un plan de redressement de la population de l’ESL, en combinant les conséquences directes et indirectes, afin de considérer globalement les effets des contaminants dans l’ESL.
En conclusion, les études sur la contamination des bélugas de l’ESL ont permis d’identifier et de quantifier de nombreux contaminants accumulés chez l’espèce et, par conséquent, de nous informer de la présence de cette contamination dans son habitat. Il serait cependant erroné de considérer le béluga comme un juste indicateur de la décontamination de son habitat. En effet, trop de facteurs et paramètres, incluant le transfert intergénérationnel des contaminants et la longévité de l’espèce, suggèrent que les tendances temporelles de la décontamination chez le béluga de l’ESL ne représentent pas nécessairement celles dans son habitat. Cependant, en raison du niveau trophique élevé occupé par le béluga dans l’ESL, ses tissus atteignent rapidement des niveaux mesurables de composés organiques persistants. Ainsi, le béluga de l’ESL doit être considéré comme une espèce sentinelle de la présence de POP émergents dans son habitat. Une analyse chimique élargie des tissus du béluga de l’ESL permettrait de déceler à un stage précoce la présence de nouveaux composés organiques persistants et bioaccumulables qui contaminent son habitat. La capacité du béluga à agir comme espèce sentinelle des contaminations émergentes dans son habitat devra être mise à profit pour une gestion préventive de la qualité de son habitat.
Bien que le manque de rétablissement de la population du béluga de l’ESL ne soit pas formellement associé à son exposition aux contaminants, on ne peut ignorer cette cause. En effet, les bélugas de l’ESL sont contaminés par des composés organiques persistants ayant démontré des effets toxiques chez d’autres organismes, incluant des perturbations au niveau de la reproduction, une augmentation des taux de mortalité, une réduction du système immunitaire ainsi que des effets comportementaux. Il est donc nécessaire de poursuivre les recherches afin de mieux comprendre l’impact des contaminations dans l’écosystème de l’ESL et sur sa population de bélugas. En somme, la précarité de la population des bélugas de l’ESL pourrait refléter la fragilité de son écosystème.
Parties annexes
Remerciements
L’auteur souhaite souligner le leadership exceptionnel de Pierre Béland (Ph.D.) et Daniel Martineau (DMV, Ph.D.) à faire connaître et à documenter depuis plus de 25 ans la problématique de la contamination et des risques toxiques inhérents auxquels est confrontée la population des bélugas de l’estuaire du Saint-Laurent.
Références bibliographiques
- AHLBORG U.G., G.C. BECKING, L.S. BIRNBAUM, A. BROUWER, H.J.G.M. DERKS M. FEELEY, G. GOLOR, A. HANBERG, J.C. LARSEN, A.K.D. LIEM, S.H. SAFE, C. SCHLATTER, F. WAERN, M. YOUNES et E. YRJÄNHEIKKI (1994). Toxic equivalency factors for dioxin-like PCBs. Chemosphere, 28, 1049-1067.
- BAILEY R. et N. ZINGER (1995). Plan de rétablissement du béluga du Saint-Laurent. Ministère des pêches et Océans et le fonds mondial pour la nature, 73 p.
- BÉLAND P., S. DeGUISE, C. GIRARD, A. LAGACÉ, D. MARTINEAU, R. MICHAUD, D.C.G. MUIR, R.J. NORSTROM, É. PELLETIER, E. RAY et L.R. SHUGART (1993). Toxic compounds and health and reproductive effects in the St. Lawrence beluga whales. J. Great Lakes Res., 19, 766-775.
- BERGMAN Å., R.J. NORSTROM, K. HARAGUCHI, H. KUROKI et P. Béland (1994). PCB and DDE methyl sulfones in mammals from Canada and Sweden. Envrion. Toxicol. Chem., 13, 121-128.
- BENNIE D.T., C.A. SULLIVAN et R.J. MAGUIRE (2000). Occurence of chlorinated paraffins in beluga whales (Delphinapterus leucas) from the St. Lawrence River and Rainbow trout and Carp from Lake Ontario. Water Qual. Res. J. Can., 35, 263-281.
- BERNT K.E., M.O. HAMMILL, M. LEBEUF et K.M. KOVACS (1999). Levels and patterns of PCBs and OC pesticides in harbour and grey seals from the St. Lawrence Estuary, Canada. Sci. Total Environ., 244, 243–262.
- BOON J.P., I. OOSTINGH, J. Van Der MEER et M.T.J. HIILEBRAND (1994). A model for the bioaccumulation of chlorobiphenyl congeners in marine mammals. Europ. J. Pharmacol., 270, 237–351.
- BOON J.P., J. Van Der MEER, C.R. ALLCHIN, R.J. LAW, J. KLUNGSOYR, P.E.G. LEONARDS, H. SPLIID, E. STROO-HANSEN, C. McKENZIE et D.E. WELLS (1997). Concentration-dependent changes of PCB patterns in fish-eating mammals: structural evidence for induction of cytochrome P450. Arch. Environ. Contam. Toxicol., 33, 298-311.
- BROUSSEAU P., S. DeGUISE, I. VOCCIA, S. RUBY et M. FOURNIER (2003). Immune status of the St. Lawrence estuary beluga whales. Dans : Toxicology of Marine Mammals, chapitre 14. VOS, J.G., G.D. BOSSART, M. FOURNIER et M. O’SHEA (Éditeurs). Taylor & Francis, New-York, USA, pp. 381-403.
- CAREY J., P. COOK, J. GIEPSY, P. HODSON, D. MUIR, J. OWENS et K. SOLOMON (Éditeurs) (1998). Ecotoxicological risk assessment of the chlorinated organic chemicals. Dans : SETAC Pellston Workshop on Environmental Risk Assessment for Organochlorine Compounds; Jul 24-29 1994, Allison, Ontario, Canada. Society of Environmental Toxicology and Chemistry (SETAC), Pensacola, Florida, USA. 397 p.
- CASTONGUAY M., P.V. HODSON, C.M. COUILLARD, M.J. ECKERSLEY, J.-D. DUTIL et G. VERREAULT (1994). Why is recruitment of the American eel declining in the St. Lawrence River and Gulf? Can. J. Fish. Aquat. Sci., 1, 479–488.
- CONVENTION DE STOCKHOLM (2001). http://www.pops.int/.
- COSEPAC, http://www.cosepac.gc.ca.
- de BOER J. (2000). Tris(4-chlorophenyl) methanol and Tris(4‑chlorophenyl) methane. Dans : New Types of Persistent Halogenated Compounds. Handbook of environmental chemistry, volume 3, partie K. PAASIVIRTA J. (Éditeur), Springer-Verlag, Berlin, Allemagne, pp. 31-42.
- DEVIGNE C. (2003). La contamination hépatique des bélugas (Delphinaterus Leucas) du Saint-Laurent et du détroit d’Hudson par les biphényles polychlorés (BPC) et les pesticides organochlorés. Mémoire de maîtrise (M.Sc.) Université du Québec à Rimouski, Canada, 144 p.
- FISK A.T., R.J. NORSTROM, C.H. CYMBALISTY et D.C.G. MUIR (1998). Dietary accumulation and depuration of hydrophobic organochlorines: bioaccumulation parameters and their relationship with the octanol/water partition coefficient. Environ. Toxicol. Chem., 17, 951-961.
- GAUTHIER J.M., É. PELLETIER, C. BROCHU, S. MOORE, C.D. METCALFE et P. BÉLAND (1998). Environmental contaminants in tissues of a neonate St. Lawrence beluga whale (Delphinapterus leucas). Mar. Poll. Bul., 36, 102-108.
- GAUTHIER L.T., C.E. HEBERT, D.V.C. WESELOH et R.J. LETCHER (2007). Current-use of flame retardants in the eggs of herring gulls (Larus argentatus) from the Laurentian Great Lakes. Environ. Sci. Technol., 41, 4561–4567.
- GEBBINK W.A., C. SONNE, R. DIETZ, M. KIRKEGAARD, E.W. BORN, D.C.G. MUIR et R.J. LETCHER (2008). Target tissue selectivity and burdens of diverse classes of brominated and chlorinated contaminants in polar bears (Ursus maritimus) from east Greenland. Environ, Sci. Technol., 42, 752-759.
- GERACI J.R. et V.J. LOUNSBURY (2005).Marine mammals ashore: a field guide for strandings. 2e édition. Baltimore, MD, National Aquarium in Baltimore, 372 p.
- GIESY J.P., S.A. MABURY, J.W. MARTIN, K. KANNAN, P.D. JONES, J.L. NEWSTED et K. COADY (2006). Perfluorinated compounds in the Great Lakes. Dans : Persistent Organic Pollutants in the Great Lakes. Handbook of environmental chemistry, volume 5, partie N. Hites R.A. (Éditeur), Springer-Verlag, Berlin, Allemagne, pp. 391‑438.
- GOUTEUX B., M. LEBEUF, D.C.G. MUIR et J.-P. GAGNÉ (2003). Levels and temporal trends of toxaphene congeners in beluga whales (Delphinapterus leucas) from the St. Lawrence estuary, Canada. Environ. Sci. Technol., 37, 4603-4609.
- GOUTEUX B., M. LEBEUF, M.O. HAMMILL, D.C.G. MUIR et J.-P. GAGNÉ (2005). Comparison of toxaphene congeners levels in five seals species from Eastern Canada: What is the importance of biological factors. Environ. Sci. Technol., 39, 1448-1454.
- HAMMILL M.O., L.N. MEASURES, J.-F., GOSSELIN et V. LESAGE (2007). Lack of recovery in the St. Lawrence Estuary beluga. Secrétariat Canadien de Consultation Scientifique (SCCS). Pêches et Océans Canada. Document de recherche 2000/26, 28 p.
- HEBERT C.E. et K.A. KEENLEYSIDE (1995).To normalize or to not normalize? Fat is the question. Environ. Toxicol. Chem., 14, 801-807.
- HELM P.A., K. KANNAN et T.F. BIDLEMAN (2006). Polychlorinated naphthalenes in the Great Lakes. Dans : Persistent Organic Pollutants in the Great Lakes. Handbook of environmental chemistry, vol. 5, part N. Hites R.A. (Éditeur), Springer-Verlag, Berlin, Allemagne, pp. 267‑273.
- HICKIE B.E., M.C.S. KINGSLEY, P.V. HODSON, D.C.G. MUIR, P. BÉLAND et D. MACKAY (2000). A modelling-based perspective on the past, present, and future polychlorinated biphenyl contamination of the St. Lawrence beluga whale (Delphinapterus leucas) population. Can. J. Fish. Aquat. Sci., 57(suppl. 1), 101-112.
- HITES R.A. (2006). Brominated flame retardants in the Great Lakes. Dans : Persistent Organic Pollutants in the Great Lakes. Handbook of environmental chemistry, vol. 5, part N. HITES R.A. (Éditeur), Springer-Verlag, Berlin, Allemagne, pp. 355-390.
- HOUDE M., P.F. HOEKSTRA, K.R. SOLOMON et D.C.G MUIR (2005). Organohalogen contaminants in delphinoid cetaceans. Rev. Environ. Contam. Toxicol., 184, 1-57.
- HOUDE M., J.W. MARTIN, R.J. LETCHER, K. SOLOMON et D.C.G. MUIR (2006). Biological monitoring of polyfluoroalkyl substances: A Review. Environ. Sci. Technol., 40, 3463-3473.
- HOBBS K.E., D.C.G. MUIR, R. MICHAUD, P. BÉLAND, R.J. LETCHER et R. NORSTROM (2003). PCBs and organochlorine pesticides in blubber biopsies from free-ranging St. Lawrence river estuary beluga whales (Delphinapterus leucas), 1994-1998. Environ. Pollut., 122, 291-302.
- JARMAN W.M., M. SIMON., R.J. NORSTROM, S.A. BURNS, C.A. BACON, B.R.T. SIMONELT et R.W. RISEBROUGH (1992). Global distribution of tris(4-chlorophenyl)methanol in high trophic level birds and mammals. Environ. Sci. Technol., 26, 1770-1774.
- JEPSON P.D., P.M.BENNETT, C.R ALLCHIN, R.J.LAW, T. KUIKEN, J.R.BAKER, E. ROGAN et J.K. KIRKWOOD (1999). Investigating potential associations between chronic exposure to polychlorinated biphenyls and infectious disease mortality in harbour porpoises from England and Whales. Sci. Total Environ., 243/244, 339-348.
- JEPSON P.D. P.M.BENNETT, R. DEAVILLE, C.R. ALLCHIN, J.R. BAKER et R.J. LAW (2005). Relationships between polychlorinated biphenyls and health status in harbor porpoises (Phocoena phocoena) stranded in the United Kingdom. Environ. Toxicol. Chem., 24, 238-248.
- KICENIUK J.W., J. HOLZBECHER et A. CHATT (1997). Extractable organohalogens in tissues of beluga whales from the Canadian Arctic and St. Lawrence estuary. Environ. Pollut., 97, 205-211.
- KUIKEN T. P.M. BENNETT, C.R. ALLCHIN, J.K. KIRKWOOD, J.R. BAKER, C.H. LOCKYER, M. J. WALTON et M.C. SHELDRICK (1994). PCBs, cause of death and body condition in harbour porpoises (Phocoena phocoena) from British waters. Aquat. Toxicol., 28, 13-28.
- LAW R.J., M. ALAEE, C.R. ALLCHIN, J.P. BOON, M. LEBEUF, P. LEPOM et G.A. STERN (2003). Levels and trends of polybrominated diphenylethers and other brominated flame retardants in wildlife. Environ. Int., 29, 757-770.
- LCPE (Loi canadienne sur la protection de l’environnement) (1993). Liste des substances d’intérêt prioritaire : rapport d’évaluation. Paraffines chlorées. Gouvernement du Canada, Environnement Canada, Santé et Bien-être social Canada, Ministre des Approvisionnements et Services Canada, 41 p.
- LEBEUF M., K. E. BERNT et M.O. HAMMILL (1999). Selective bioaccumulation of PCBs by phocids from the Estuary and Gulf of St. Lawrence. Dans : Proceedings of the 19th International Symposium on Halogenated Environmental Organic Pollutants and POPs, DIOXIN 1999. Organohalogen Compd., 42, 191-195.
- LEBEUF M., K.E. BERNT, M. HAMMILL et L. MEASURES (2000). Stratification of PCBs in the blubber of beluga whales (Delphinapterus Leucas) from the St. Lawrence Estuary. Dans : Proceedings of the 20th International Symposium on Halogenated Environmental Organic Pollutants and POPs, DIOXIN 2000. Organohalogen Compd., 46, 487-490.
- LEBEUF M., K.E. BERNT, S. TROTTIER, M. NOEL, M.O. HAMMILL et L. MEASURES (2001) Tris(4-chlorophenyl)methane and tris(4-chlorophenyl)methanol in marine mammals from the Estuary and Gulf of St. Lawrence. Environ. Pollut., 111, 29-43.
- LEBEUF M., B. GOUTEUX, L. MEASURES et S. TROTTIER (2004). Levels and temporal trends (1988-1999) of polybrominated diphenyl ethers in beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Canada. Environ. Sci. Technol., 38, 2971-2977.
- LEBEUF M. et T. NUNES (2005). PCBs and OCPs in sediment cores from the Lower St. Lawrence Estuary, Canada: Evidence of fluvial inputs and time lag in delivery to coring sites. Environ Sci. Technol., 39, 1470-1478.
- LEBEUF M., H. FROUIN, M. NOËL, S. TROTTIER, L. MEASURES et M. HAMMILL (2005). Distribution des contaminants persistants dans la panicule adipeuse chez le béluga du Saint-Laurent. Dans : 73e Congrès de l’ACFAS. 9-13 mai, Chicoutimi, Québec, Canada.
- LEBEUF M., C.M. COUILLARD, B. LÉGARÉ et S. TROTTIER (2006). Effects of DeBDE and PCB-126 on hepatic concentrations of PBDEs and methoxy-PBDEs in Atlantic Tomcod. Environ. Sci. Technol., 40, 3211-3216.
- LEBEUF, M., M. NOËL, S. TROTTIER et L. MEASURES (2007a). Temporal trends (1987-2002) of persistent, bioaccumulative and toxic (PBT) chemicals in beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Canada. Sci. Total. Environ., 383, 216-231.
- LEBEUF M., S. TROTTIER, M. NOËL et M.O. HAMMILL (2007b). Selectivity in the lactational transfer of PBDEs by hooded seals from the Gulf of St. Lawrence, Canada. Dans : Proceedings of 4th International Workshop on Brominated Flame Retardants, Amsterdam, Pays-Bas, 24-27 avril, BFR2007, 1-4.
- LESAGE V., M.O. HAMMILL et K.M. KOVACS (2001). Marine mammals and the community structure of the Estuary and Gulf of St. Lawrence, Canada: evidence from stable isotope analysis. Mar. Ecol. Prog. Ser., 210, 203‑221.
- LETCHER R.J., E. KLASSON-WEHBER et Å. BERGMAN (2000a). Methyl sulfone and hydroxylated metabolites of polychlorinated biphenyls. Dans : New Types of Persistent Halogenated Compounds. Handbook of environmental chemistry, volume 3, partie K. PAASIVIRTA J. (Éditeur), Springer-Verlag, Berlin, Allemagne pp. 315-359.
- LETCHER R.J., R.J. NORSTROM, D.C.G. MUIR, C.D. SANDAU, K. KOCZANSKI, R. MICHAUD, S. DeGUISE et P. BÉLAND (2000b). Methylsulfone polychlorinated biphenyl and 2,2-Bis(Chlorophenyl)-1,1-Dichloroethylene metabolites in beluga whale (Delphinapterus leucas) from the St. Lawrence River Estuary and Western Hudson Bay, Canada. Environ. Toxicol. Chem., 19, 1378-1388.
- MARTEL L.M., J. GAGNON, R. MASSÉ, A. LECLERC et L. TREMBLAY (1986). Polyclyclic aromatic hydrocarbons in sediments from the Saguenay fjord, Canada. Bull. Environ. Contam. Toxicol., 37, 133‑140.
- MARTINEAU D., P. BÉLAND, C. DESJARDINS et A. LAGACÉ (1987). Levels of organochlorine chemicals in tissues of beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Québec, Canada. Arch. Environ. Contam. Toxicol., 16, 137-147.
- MARTINEAU D., K. LEMBERGER, A. DALLAIRE, P. LABELLE, T.P. LIPSCOMB, P. MICHEL et I. MIKAELIAN (2002). Cancer in wildlife, a case study: Beluga from the St. Lawrence estuary, Que´bec, Canada. Environ. Health Perspec., 110, 285-292.
- MASSÉ, R., D. MARTINEAU, L.TREMBLAY et P. BÉLAND (1986). Concentrations and chromatographic profile of DDT metabolites and polychlorobiphenyl (PCB) residues in stranded Beluga Whales (Delphinapterus leucas) from the St. Lawrence Estuary, Canada. Arch. Environ. Contam. Toxicol., 15, 567-579.
- McALPINE, D.F., M.C.S. KINGSLEY, P.-Y. DAOUST (1999). A lactating record-age St. Lawrence beluga (Delphinapterus leucas). Mar. Mam. Sci., 15, 854-859.
- McKINNEY, M.A., S. DeGUISE, D. MARTINEAU, P. BÉLAND, M. LEBEUF et R.J. LETCHER (2006). Organohalogen contaminants and metabolites in beluga whale (Delphinapterus leucas) liver from two Canadian populations. Environ. Toxicol. Chem., 25, 1246-1257.
- MIKAELIAN, I., C.W.Jr. QUALLS, S. DeGUISE, M.W. WHALEY et D. MARTINEAU (1999). Bone fluoride concentrations in beluga whales from Canada. J. Wildlife Dis., 35, 356-360.
- METCALFE C., T. METCALFE, S. RAY, G. PATERSON et B. KOENIG (1999). Polychlorinated biphenyls and organochlorine compounds in brain, liver and muscle of beluga whales (Delphinapterus leucas) from the Arctic and St. Lawrence estuary. Mar. Environ. Res., 47, 1-15.
- MICHAUD R. (1993). Distribution estivale du béluga du Saint-Laurent; synthèse 1986 à 1992. Rapp. Tech. Can. Sci. Halieut. Aquat., Document 1906, 28 p.
- MEASURES L. (2007). Compte rendu de l’atelier de travail sur le béluga de l’estuaire du Saint-Laurent – revue de programme de suivi des carcasses. Secrétariat Canadien de Consultation Scientifique (SCCS). Pêches et Océans Canada. Compte rendu. 2007/05, 94 p.
- MEASURES L. (2008). Les causes de mortalité du béluga du Saint-Laurent. Nat. Can., 132, 75-79.
- MUIR D.C.G., C.A. FORD, R.E.A. STEWART, T.G. SMITH, R.F. ADDISON, M.E. ZINCK et P. BÉLAND (1990). Organochlorine contaminants in belugas (Delphinapterus leucas) from Canadian waters. Can. Bull. Fish. Aquat. Sci., 224, 165-190.
- MUIR, D.C.G., C.A. FORD, B. ROSENBERG, R.J. NORSTROM, M. SIMON et P. BÉLAND (1996a). Persistent organochlorines in beluga whales (Delphinapterus leucas) from the St. Lawrence River Estuary-I. Concentrations and patterns of specific PCBs, chlorinated pesticides and polychlorinated dibenzo-p-dioxins and dibenzofurans. Environ. Poll., 93, 219-234.
- MUIR, D.C. G., K. KOCZANSKI, R. ROSENBERG et P. BÉLAND (1996b). Persistent organochlorines in beluga whales (Delphinapterus leucas) from the St. Lawrence River Estuary - II. Temporal trends, 1982-1994. Environ. Poll., 93, 235-245.
- MYERS R.A., J.A. HUTCHINGS et N.J. BARROWMAN (1997). Why do fish stocks collapse? The example of cod in Atlantic Canada. Ecol. Appl., 7, 91‑106.
- NATO-CCMS (NORTH ATLANTIC TREATY ORGANIZATION - COMMITTEE ON THE CHALLENGES OF MODERN SOCIETY)(1988). Pilot study on international information exchange on dioxins and related compounds: International toxicity equivalency factor (ITEF). Method of risk assessment for complex mixture of dioxins and related compounds. Rep. No. 176/178, ?? p.
- NIIMI A.J., C.D. METCALFE et S.Y. HUESTIS (1994). Chlorinated diphenylethers in Great Lakes fish and their environmental implication. Environ. Toxicol. Chem., 13, 1133-1138.
- NORSTROM, R.J., D.C.G. MUIR, C.A. FORD, M. SIMON, C.R. MACDONALD et P. BÉLAND (1992). Indications of P450 monooxygenase activities in beluga (Delphinapterus leucas) and narwhal (Monodon monoceros) from patterns of PCB, PCDD and PCDF accumulation. Mar. Envrion. Res., 34, 267-272.
- PMSSL (Parc marin Saguenay-Saint-Laurent), http://www.parcmarin.qc.ca/1508_fr.html.
- ROSS P.S., C.M. COUILLARD, M.G. IKONOMOU, S.C. JOHANNESSEN, M. LEBEUF, R.W. MACDONALD et G. TOMY (2009). Large and growing environmental reservoirs of Deca-BDE present an emerging health risk for fish and marine mammals. Mar. Pollut. Bull., 58, 7-10.
- SERGEANT D.E. (1980). Levels of mercury and organochlorine residues in tissues of sea mammals from the St. Lawrence estuary. Conseil International pour l’Exploration de la Mer (CIEM) C.M. /E:55.
- STAPLETON H.M., R.J. LETCHER et J.E. BAKER (2004). Debromination of polybrominated diphenyl ether congeners BDE99 and BDE 183 in the intestinal tract of the common carp (Cyprinus carpio). Environ. Sci. Technol., 38, 1054-1061.
- STEWART R.E.A., S.E. CAMPANA, C.M. JONES et B.M. STEWART (2007). Bomb radiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates. Can. J. of Zool., 84,1840-1852.
- ST-LOUIS R., S. DE MORA, É. PELLETIER, D. DOIDGE, D. LECLAIR, I. MIKAELIAN et D. MARTINEAU (2000). Hepatic butyltin concentrations in beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary and Northern Quebec. Appl. Organometal. Chem., 14, 218‑226.
- TANABE S, S. WATANABE, H. KAN et R. TATSUKAWA (1988). Capacity and mode of metabolism in small cetaceans. Mar. Mamm. Sci., 4,103‑124.
- TITTLEMIER, S. A., A. BORRELL, J. DUFFE, P.J. DUIGNAN, P. FAIR, P. HOEKSTRA, K.M. KOVACS, M.M. KRAHN, M. LEBEUF, C. LYDERSEN, D. MUIR, T. O’HARA, M. OLSSON, J. PRANSCHKE, P. ROSS, U. SIEBERT, G. STERN, S. TANABE et R. NORSTROM (2002). Global distribution of halogenated dimethyl bipyrroles in marine mammals blubber. Environ. Contam. and Toxicol., 43, 244-255.
- TOMY G.T., D.C.G. MUIR, G. STERN et J.B. WESTMORE (2000). Levels of C10_C13 polychloro-n-alkanes in marine mammals from arctic and the St. Lawrence river estuary. Environ. Sci. Technol., 34, 1615-1619.
- TEUTEN E.L., L. XU et C.M. REDDY (2005). Two abundant bioaccumulated halogenated compounds are natural products. Science, 307, 917-920.
- Van Den BERG, M., L.S. BIRNBAUM, M. DENISON, M. DE VITOB, W. FARLAND, M. FEELEY, H. FIEDLER, H. HAKANSSON, A. HANBERG, L. HAWS, M. ROSE, S. SAFE, D. SCHRENK, C. TOHYAMA, A. TRITSCHER, J. TUOMISTO, M. TYSKLIND, N. WALKER et R.E. PETERSON (2006). The 2005 World Health Organization reevaluation of human and mammalian toxic equivalency factors for dioxins and dioxin-like compounds. Toxicol. Sci., 93, 223‑241.
- WAGEMANN R., R.E.A. STEWART. P. BÉLAND et C. DESJARDINS (1990). Heavy metals and selenium in tissues of belugas whales (Delphinapterus leucas) from the Canadian Arctic and the St. Lawrence Estuary. Can. Bull. Fish. Aquat. Sci., 224,191-206.
- YANG F., Y.K. CHAU et R.J. MAGUIRE (1998). Occurence of butyltin compounds in beluga whales (Delphinapterus leucas). Appl. Organometal. Chem., 12, 651-656.
- ZPM Estuaire, zone de protection marine de l’estuaire du Saint-Laurent, http://www.qc.dfo-mpo.gc.ca/ZPMEstuaire/fr/proj.asp.
Liste des figures
Figure 1
Structure chimique générale des biphényles polychlorés (BPC).
General chemical structure of polychlorinated biphenyls (PCB).
Figure 2
Structures chimiques du a) 4, 4’DDT et b) 4, 4’DDE.
Chemical structures of a) 4, 4’DDT and b) 4, 4’DDE.
(a)
(b)
Figure 3
Structures chimiques des congénères de toxaphène a) P26 et b) P50.
Chemical structures of toxaphene congeners a) P26 and b) P50.
(a)
(b)
Figure 4
Structures chimiques du a) γ-Chlordane, b) Lindane (γ-HCH), c) hexachlorobenzène (HCB), d) Mirex et e) tris-(4-chlorophényle) méthanol (TCPMOH).
Chemical structures of a) γ-Chlordane, b) Lindane (γ-HCH), c) hexachlorobenzene (HCB), d) Mirex and e) tris-(4‑chlorophenyl) methanol (TCPMOH).
(a)
(b)
(c)
(d)
(e)
Figure 5
Structures chimiques générales des a) polychlorodibenzo-p-dioxine (PCDD) et b) polychlorodibenzofuranne (PCDF).
General chemical structures of a) polychlorinated dibenzo-p-dioxin (PCDD) and b) polychlorinated dibenzofuran (PCDF).
(a)
(b)
Figure 6
Structures chimiques du a) polychlo-n-alkane (PCA) (C10), b) dieldrine, c) endrine et d) octachlorostyrène.
Chemical structures of a) polychloro- n-alkanes (PCA) (C10), b) dieldrin, c) endrin and d) octachlorostyrene.
(a)
(b)
(c)
(d)
Figure 7
Structure chimique générale des polybro-modiphényles éthers (PBDE).
General chemical structure of polybrominated diphenylethers (PBDE).
Liste des tableaux
Tableau 1
Concentrations des biphényles polychlorés, BPC, (moyenne ± écart-type, moyenne géométrique et/ou gamme) et contribution relative des groupes homologues (%) dans les tissus des bélugas du Saint-Laurent
Concentrations of polychlorinated biphenyls, PCB, (mean ± std dev, geometric mean and/or range) and relative contribution of homolog groups (%) in tissues of the St. Lawrence belugas
|
|
|
|
|
Moyenne ± écart-type (moyenne géométrique et/ou gamme) |
Contribution des groupes homoloques à la somme arithmétique des BPC ± écart-type (ou à la somme géométrique des BPC) |
|
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
BPC Aroclors (µg•g‑1 lip) |
ΣBPC congénères (µg•g‑1 lip) |
Mono- Tri (%) |
Tétra (%) |
Penta (%) |
Hexa (%) |
Hepta (%) |
Octa-Déca (%) |
Référence |
Gras |
C |
F2 |
(juvénile-adulte) |
1971-1979 |
486 (171-800) |
|
|
|
|
|
|
|
SERGEANT et al., 1980 |
|
C |
M2 |
39 (33-44) |
1983-1984 |
68,4 (68,0) (61,6- 75,2) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
|
C |
M15 |
34 (<1-55) |
1982-1985 |
210 ± 98 (187) (77,7- 344) |
|
|
|
|
|
|
|
MARTINEAU et al., 1987 |
|
C |
F10 |
40 (3,5 -59) |
1982-1985 |
117 ± 211 (50,0) (17,7 – 708) |
|
|
|
|
|
|
|
Idem |
|
C |
M4 |
35 (8-47) |
1986-1987 |
137,1±29,4 (96,3-165,9) |
87,3 ± 17,6 (62,1-103) |
< 0,1 |
18,3 ±1,3 |
11,1 ±1,9 |
36,6 ±8,2 |
26,9 ±8,2 |
7,1 ±3,8 |
MUIR et al., 1990 |
|
C |
F5 |
31 (5-58) |
1986-1987 |
62,6±43,4 (18,9-129,3) |
43,1 ± 25,4 (16,7- 79,3) |
< 0,1 |
15,7 ±12,2 |
14,7 ±6,8 |
34,0 ±22,0 |
27,6 ±16,9 |
8,0 ± 4,8 |
Idem |
|
C |
?3 |
? |
1952- 1986 |
16,6 ± 19,6 (9,5) (2,7-39,0) |
|
|
|
|
|
|
|
JARMAN et al., 1992 |
|
C |
F1 |
50 |
1988 |
|
22,6 |
|
|
|
|
|
|
BERGMAN et al., 1994 |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
(86,0) (7,9-451) |
112,4 ± 98,8 (78,9) (8,3 – 412) |
(< 0,1) |
(13,2) |
(22,7) |
(42,5) |
(18,5) |
(3,2) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987- 1990 |
(30,1) (7,3-90,7) |
35,2 ± 21,0 (29,6) (8,8 – 83,3) |
(0,1) |
(8,5) |
(22,0) |
(39,1) |
(23,9) |
(6,4) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
(97,2) (54,8-145) |
77,3 ± 23,1 (74,5) (48,2 – 126) |
(0,4) |
(9,3) |
(20,4) |
(46,0) |
(19,8) |
(4,1) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
(69,1) (13,5-174) |
77,3 ± 54,9 (57,8) (14,3 – 174) |
(0,3) |
(8,1) |
(20,4) |
(44,2) |
(22,8) |
(4,2) |
Idem |
|
C |
F1 |
68 |
1996 |
|
9,4 |
|
|
|
|
|
|
McALPINE et al., 1999 |
|
C |
?1 |
<1 |
1991 |
|
17,6 |
nd |
13,4 |
17,3 |
50,7 |
17,3 |
1,2 |
GAUTHIER et al., 1998 |
|
B |
M30 |
14* (8-22) |
1994-1996 |
|
7,5 (2,0 - 86,3) |
|
|
|
|
|
|
LETCHER et al., 2000b |
|
B |
F3 |
12* (7-15) |
1994-1996 |
|
10,9 (8,5 – 19,7) |
|
|
|
|
|
|
Idem |
|
B |
M34 |
14* (8-22) |
1994-1998 |
|
20,6 ± 26,8 (12,2) (2,1 – 128) |
(20,3) |
(10,8) |
(19,5) |
(29,0) |
(16,8) |
(3,5) |
HOBBS et al., 2003 |
|
B |
F10 |
14* (7-22) |
1994-1998 |
|
21,2 ± 14,3 (11,8) (0,15- 44,1) |
(8,4) |
(13,2) |
(25,0) |
(38,1) |
(12,7) |
(2,60) |
Idem |
|
C |
M44 |
42 (22-74) |
1988-2002 |
|
85,3 ± 39,5 (75,0) (5,4 – 209) |
0,03 (6,0) |
9,1 (17,9) |
14,8 (19,0) |
49,0 (21,4) |
23,7 (19,9) |
3,3 (15,8) |
LEBEUF et al., 2007a |
|
C |
F42 |
40 (20-64) |
1987-2002 |
|
20,2 ± 17,2 (15,3) (2,6 – 83,1) |
0,13 (7,7) |
6,9 (16,3) |
16,3 (18,7) |
42,4 (20,9) |
28,3 (20,0) |
5,9 (16,4) |
Idem |
Foie |
C |
M3 |
36 (30-44) |
1983-1984 |
5,4 ± 4,9 (4,2) (2,3-11,1) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
|
C |
M10 |
31 (<1-48) |
1983-1985 |
262±153 (233) (61,5-486) |
|
|
|
|
|
|
|
MARTINEAU et al., 1987 |
|
C |
F7 |
37 (3,5 -56) |
1983-1985 |
653±1323 (82,4) (5,5-3869) |
|
|
|
|
|
|
|
Idem |
|
C |
?1 |
<1 |
1991 |
|
8,8 |
|
|
|
|
|
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
53,9 (18,4-81,1) |
0,6 |
12,7 |
18,1 |
46,9 |
14,3 |
7,9 |
METCALFE et al., 1999 |
|
C |
M11 |
21 (<1-48) |
1995-1999 |
|
46,3±39,2 (33,6) (9,3-125) |
|
|
|
|
|
|
DEVIGNE 2003 |
|
C |
F13 |
32(12) (<1-62) |
1995-1999 |
|
39,5±79,5 (12,9) (1,5-293) |
|
|
|
|
|
|
Idem |
|
C |
M3/ F3 |
40 (11-55) 5,3 (<1-9) |
2000-2003 |
|
31,9±6,5 (1,4-45,4) |
< 0,1 |
15,5 |
20,3 |
35,8 |
23,0 |
5,3 |
McKINNEY et al., 2006 |
Rein |
C |
M3 |
36 (30-44) |
1983-1984 |
19 ± 27 (8,6) (3,4-50,2) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
|
C |
M9 |
30 (<1-48) |
1983-1985 |
135±51,4 (124) (57,0-222) |
|
|
|
|
|
|
|
MARTINEAU et al., 1987 |
|
C |
F7 |
37 (3,5 -56) |
1983-1985 |
454±700 (76,2) (4,5-1930) |
|
|
|
|
|
|
|
Idem |
|
C |
?1 |
<1 |
1991 |
|
4,1 |
|
|
|
|
|
|
GAUTHIER et al., 1998 |
Poumon |
C |
M2 |
39 (33-44) |
1983-1984 |
4,4 (4,4) (4,2-4,7) |
|
|
|
|
|
|
|
MASSE et al., 1986 |
Lait |
C |
F1 |
46 |
1984 |
3,8 |
|
|
|
|
|
|
|
MASSE et al., 1986 |
Cerveau |
C |
?1 |
<1 |
1991 |
|
1,7 |
|
|
|
|
|
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
42,9 (8,5-111) |
1,0 |
14,7 |
17,2 |
46,1 |
14,6 |
6,6 |
METCALFE et al., 1999 |
Muscle |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
64,8 (41,2-105) |
0,6 |
13,8 |
15,7 |
48,4 |
14,7 |
6,8 |
METCALFE et al., 1999 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal.
Tableau 2
Concentrations du dichlorodiphényltrichloroéthane, DDT, et ses métabolites (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas du Saint-Laurent*.
Concentrations of dichlorodiphenyltrichloroethane, DDT, and its metabolites (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
Moyenne (µg•g‑1 lipide) ± écart-type (moyenne géométrique et/ou gamme) |
Ratio |
|
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
Sexe, N |
Années |
4, 4’DDT |
4, 4’DDE |
4, 4’DDD |
2, 4’DDT |
2, 4’DDE |
2, 4’DDD |
ΣDDT |
4, 4’DDE /ΣDDT |
Référence |
Gras |
F2 |
1971-1979 |
132 (1,5-262) |
231 (24,9-437) |
(<0,01-128) |
|
|
|
431 (34-827) |
0,63 (0,53-0,73) |
SERGEANT et al., 1980 |
|
M2 |
1983-1984 |
|
81,0 (80,1) (69,0-93,0) |
|
|
|
|
93,7 (92,5) (79,3-108) |
0,87 (0,86) (0,86-0,87) |
MASSE et al., 1986 |
|
M15 |
1982-1985 |
22,4±19,6 (11,0) (0,3-66,1) |
57,9±37,8 (45,9) (8,6-150) |
17,3±10,3 (14,0) (3,2-39,4) |
|
|
|
97,6±64,6 (76,0) (14,5-248) |
0,61±0,07 (0,60) (0,50-0,80) |
MARTINEAU et al., 1987 |
|
F10 |
1982-1985 |
6,8±6,6 (3,9) (0,9-17,3) |
11,4±19,3 (4,2) (1,1-65,1) |
6,1±9,7 (2,6) (0,4-34,2) |
|
|
|
27,2±30,0 (14,8) (2,5-117) |
0,36±0,17 (0,28) (0,05-0,56) |
Idem |
|
M4 |
1986-1987 |
|
75,8±22,0 (44,4-92,9) |
|
|
|
|
116,4±37,6 (60,4-142) |
0,66±0,06 |
MUIR et al., 1990 |
|
F5 |
1986-1987 |
|
16,1±13,0 (2,0-30,9) |
|
|
|
|
26,6±20,0 (4,6-49,3) |
0,56±0,09 |
Idem |
|
?3 |
1952- 1986 |
|
|
|
|
|
|
38,1±39,1 (21,8) (4,4-81,0) |
|
JARMAN et al., 1992 |
|
F1 |
1988 |
|
|
|
|
|
|
16,6 |
|
BERGMAN et al., 1994 |
|
M15 |
1988- 1990 |
(13,6) (0,4-84,3) |
(47,1) (2,1-249) |
(8,6) (0,8-39,2) |
(4,1) (0,05-16,6) |
|
|
120±103 (76,0) (3,4-389) |
0,62 |
MUIR et al., 1996a |
|
F21 |
1987- 1990 |
(3,6) (1,0-15,4) |
(10,4) (1,7-52,3) |
(2,6) (0,6-9,1) |
(0,66) (0,17-3,98) |
|
|
24,3±18,6 (18,6) (3,6-80,8) |
0,56 |
Idem |
|
M9 |
1993-1994 |
(9,3) (1,7-17,8) |
(24,9) (12,6-31,5) |
(6,6) (1,3-11,7) |
(4,4) (0,9-8,5) |
(0,36) (0,16-0,74) |
(0,26) ((0,11-0,45) |
51,5±16,0 (48,4) (20,5-64,6) |
0,51 |
MUIR et al., 1996b |
|
F7 |
1993-1994 |
(7,1) (1,1-20,3) |
(16,6) (3,3-37,5) |
(5,5) (0,9-13,6) |
(2,5) (0,3-9,6) |
(0,23) (0,05-0,51) |
(0,42) (0,29-1,22) |
45,1±27,8 (32,9) (5,7-77,6) |
0,50 |
Idem |
|
F1 |
1996 |
|
|
|
|
|
|
7,3 |
|
McALPINE et al., 1999 |
|
?1 |
1991 |
0,11 |
2,1 |
0,02 |
|
|
|
2,2 |
0,95 |
GAUTHIER et al., 1998 |
|
M30 |
1994-1996 |
|
3,7 (0,3-41,7) |
|
|
0,39 (0,03-4,5) |
|
|
|
LETCHER et al., 2000b |
|
F3 |
1994-1996 |
|
1,5 (0,6-1,8) |
|
|
0,36 (0,3-0,76) |
|
|
|
idem |
|
M5 |
1997-1998 |
|
83,8 (15,5-125) |
|
|
|
|
127 (25,1-190) |
0,66 |
LEBEUF et al., 2001 |
|
F5 |
1997 |
|
6,0 (1,3-9,3) |
|
|
|
|
11,7 (7,1-18,6) |
0,51 |
idem |
|
M34 |
1994-1998 |
(0,94) (0,11-30,2) |
(4,6) (0,4-138) |
(0,97) (0,13-16,9) |
(0,64) (0,07-21,7) |
(0,43) (0,03-4,8) |
(0,39) (<0,01-4,0) |
20,7±36,8 (13,3) (1,3-201) |
0,35 |
HOBBS et al., 2003 |
|
F10 |
1994-1998 |
(0,48) (<0,01-4,6) |
(1,4) (0,05-15,2) |
(0,47) (0,02-2,6) |
(0,23) (<0,01-1,8) |
(0,14) (<0,01-0,8) |
(0,01) (<0,01-4,8) |
7,0±7,3 (5,3) (0,09-24,4) |
0,26 |
idem |
|
M44 |
1988-2002 |
12,6±7,2 (10,6) (0,3-34,4) |
86,9±42,9 (75,8) (2,1-219) |
14,5±7,5 (13,0) (0,7-41,8) |
|
0,30±0,18 (0,25) (0,02-0,81) |
0,31±0,36 (0,25) (0,04-2,5) |
115±55,9 (101) (3,1-297) |
0,76±0,05 (0,75) (0,65-0,88) |
LEBEUF et al., 2007a |
|
F42 |
1987-2002 |
2,2±2,2 (1,4) (0,2-8,9) |
7,1±8,6 (4,3) (0,5-43,0) |
2,2±2,3 (1,4) (0,2-9,6) |
|
0,04±0,06 (0,03) (<0,01-0,4) |
0,08±0,06 (0,06) (0,01-0,3) |
11,6±13,0 (7,3) (0,8-61,2) |
0,60±0,05 (0,59) (0,48-0,71) |
idem |
Foie |
M3 |
1983-1984 |
|
5,4±4,6 (4,0) (1,4-10,5) |
|
|
|
|
5,8±4,8 (4,3) (1,5-11,0) |
0,94±0,02 (0,94) (0,92-0,95) |
MASSE et al., 1986 |
|
M10 |
1983-1985 |
1,9±2,0 (1,4) (<0,01-6,1) |
57,9±36,2 (47,7) (17,5-140) |
32,1±24,8 (23,9) (7,5-85,1) |
|
|
|
91,9±61,7 (74,1) (27,9-232) |
0,65±0,05 (0,64) (0,56-0,71) |
MARTINEAU et al., 1987 |
|
F7 |
1983-1985 |
2,1±1,6 (1,6) (0,3-5,7) |
45,6±101 (6,1) (0,6-293) |
21,4±45,7 (3,7) (0,5-133) |
|
|
|
69,1±148 (12,7) (2,2-432) |
0,52±0,17 (0,48) (0,17-0,70) |
idem |
|
?1 |
1991 |
0,08 |
3,4 |
0,02 |
|
|
|
3,5 |
0,97 |
GAUTHIER et al., 1998 |
|
M4 |
1988-1989 |
5,1 |
48,5 |
9,8 |
|
|
|
63,4 (15,3-75,6) |
0,77 |
METCALFE et al., 1999 |
|
M11 |
1995-1999 |
0,26±0,29 (0,09) (<0,01-0,78) |
24,7±30,4 (12,6) (2,4-94,9) |
6,7±9,2 (3,9) (1,0-32,8) |
|
|
|
31,8±39,1 (17,0) (3,5-128) |
0,74±0,08 (0,74) (0,56-0,87) |
DEVIGNE 2003 |
|
F13 |
1995-1999 |
0,18±0,31 (0,05) (<0,01-1,14) |
22,3±49,0 (3,7) (0,3-163) |
6,1±12,5 (1,4) (0,09-45,5) |
|
|
|
28,6±61,3 (5,4) (0,5-209) |
0,70±0,09 (0,70) (0,53-0,85) |
Idem |
|
M3/ F3 |
2000-2003 |
|
|
|
|
|
|
4,5±1,2 (0,04-7,8) |
|
McKINNEY et al., 2006 |
Rein |
M3 |
1983-1984 |
|
6,7±5,9 (5,1) (2,2-13,4) |
|
|
|
|
7,9±7,9 (5,6) (2,2-16,9) |
0,91±0,10 (0,91) (0,79-0,98) |
MASSE et al., 1986 |
|
M9 |
1983-1985 |
6,0±8,6 (1,7) (0,1-28,0) |
30,8±16,7 (26,4) (11,0-61,6) |
13,4±6,5 (11,4) (3,7-23,0) |
|
|
|
50,2±28,9 (41,9) (18,7-105) |
0,44±0,02 (0,44) (0,41-0,47) |
MARTINEAU et al., 1987 |
|
F7 |
1983-1985 |
3,4±3,2 (2,1) (0,4-10,4) |
40,8±56,9 (8,9) (1,1-147) |
21,0±29,5 (4,9) (0,5-78,9) |
|
|
|
65,3±87,1 (17,0) (2,0-211) |
0,51±0,03 (0,51) (0,45-0,55) |
Idem |
|
?1 |
1991 |
0,04 |
2,3 |
<0,01 |
|
|
|
2,3 |
0,98 |
GAUTHIER et al., 1998 |
Poumon |
M2 |
1983-1984 |
|
6,3 (6,2) (6,0-6,5) |
|
|
|
|
6,6 (6,6) (6,4-6,8) |
0,95 (0,94) (0,94-0,95) |
MASSE et al., 1986 |
Lait |
F1 |
1984 |
|
4,2 |
|
|
|
|
4,5 |
0,92 |
MASSE et al., 1986 |
Cerveau |
?1 |
1991 |
0,01 |
0,69 |
<0,01 |
|
|
|
0,70 |
0,98 |
GAUTHIER et al., 1998 |
|
M4 |
1988-1989 |
27,0 |
86,9 |
6,6 |
|
|
|
121 (9,3-387) |
0,72 |
METCALFE et al., 1999 |
Muscle |
M4 |
1988-1989 |
10,3 |
68,2 |
15,7 |
|
|
|
94,2 (53,7-150) |
0,72 |
METCALFE et al., 1999 |
nombre,
mâle,
femelle,
information sur l’état et l’âge des animaux au Tableau 1
Tableau 3
Concentrations du toxaphène total et de certains congénères spécifiques (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans le gras des bélugas du Saint-Laurent.
Concentrations of total toxaphene and selected congeners (mean ± std dev, geometric mean and/or range) in blubber of the St. Lawrence belugas.
|
|
|
|
|
Moyenne (µg•g‑1 lipide) ± écart-type (moyenne géométrique et/ou gamme) |
|
|||
|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
Toxaphène total |
Somme de six congénères |
P26 (T2) |
P50 (T12) |
Référence |
Gras |
C |
M4 |
35 (8-47) |
1986-1987 |
16.9±2.8 (13.5-20.3) |
|
|
|
MUIR et al., 1990 |
|
C |
F5 |
31 (5-58) |
1986-1987 |
7.3±4.1 (2.4-11.9) |
|
|
|
Idem |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
(18.0) (2.4-45.6) |
|
(1,3) (0,20-4,3) |
(4,3) (0,64-16,6) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987- 1990 |
(8.6) (3.2-2.3) |
|
(0,45) (0,17-6,1) |
(1,5) (0,37-22,3) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
(27.0) (16.9-35.9) |
|
(0,80) (0,49-1,1) |
(1,6) (1,1-2,2) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
(20.8) (6.3-40.0) |
|
(0,50) (0,12-1,0) |
(1,3) (0,73-2,4) |
Idem |
|
B |
M30 |
14* (8-22) |
1994-1996 |
(8.0) (1.0-89.7) |
|
|
|
HOBBS et al., 2003 |
|
B |
F5 |
14* (7-22) |
1994-1996 |
(2.4) (0.28-6.5) |
|
|
|
Idem |
|
C |
M26 |
41 (22-58) |
1988-1999 |
|
2,6±0,8 (1,0-4,9) |
0,80±0,24 (0,35-1,3) |
1,7±0,6 (0,61-3,5) |
GOUTEUX et al., 2003 |
|
C |
F26 |
42 (20-63) |
1987-1999 |
|
1,0±0,8 (0,22-3,3) |
0,31±0,27 (0,05-1,2) |
0,57±0,48 (0,16-2,0) |
Idem |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal
Tableau 4
Concentrations du chlordane, hexachlorocyclohexane, hexachlorobenzène, mirex et tris(4-chlorophényle) méthane/méthanol (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas du Saint-Laurent.
Concentrations of chlordane, hexachlorocyclohexane, hexachlorbenzene, mirex and tris(4-chlorophenyl) methane/methanol (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (moyenne géométrique et/ou gamme) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
CHL (µg.g‑1 lip) |
HCH (µg•g‑1 lip) |
HCB (µg•g‑1 lip) |
Mirex (µg•g‑1 lip) |
TCPM (ng•g‑1 lip) |
Référence |
Gras |
C |
M4 |
35 (8-47) |
1986-1987 |
8,6±0,7 (7,5-9,2) |
0,43±0,13 (0,32-0,59) |
1,5±0,50 (0,94-2,2) |
1,2±0,74 (0,22-1,8) |
|
MUIR et al., 1990* |
|
C |
F5 |
31 (5-58) |
1986-1987 |
4,1±2,3 (1,7-7,4) |
0,28±0,12 (0,14-0,37) |
0,69±0,50 (0,25-1,5) |
1,3±0,11 (0,44-3,1) |
|
Idem |
|
C |
?3 |
nd |
1952- 1986 |
|
|
|
|
15±22 (1-41) |
JARMAN et al., 1992 |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
(8,4) (1,4-28) |
(0,47) (0,20-0,90) |
(1,4) (0,24-5,5) |
(0,33) (<0,01-6,8) |
(434) (45-1783) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987- 1990 |
(3,5) (1,2-10) |
(0,29) (0,11-0,77) |
(0,37) (0,08-1,8) |
(1,1) (0,18-5,0) |
(274) (111-701) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
(11) (8,2-16) |
(0,35) (0,28-0,51) |
(0,97) (0,48-1,9) |
(1,0) (0,66-1,9) |
(409) (220-1044) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
(8,9) (2,3-18) |
(0,30) (0,18-0,56) |
(0,67) (0,11-1,7) |
(0,90) (0,18-2,4) |
(336) (220-832) |
Idem |
|
C |
?1 |
<1 |
1991 |
1,4 |
0,16 |
0,28 |
0,06 |
|
GAUTHIER et al., 1998 |
|
C |
M5 |
38 (24-54) |
1997-1998 |
|
|
|
|
34 (8,8-50) |
LEBEUF et al., 2001 |
|
C |
F5 |
57 (42-62) |
1997 |
|
|
|
|
13 (3,5-18) |
Idem |
|
B |
M30 |
14* (8-22) |
1994-1998 |
(2,0) (0,25-8,1) |
(0,53) (0,05-3,6) |
(0,18) (0,04-0,85) |
(0,12) (0,02-1,6) |
(3,1) (2,1-9,0) |
HOBBS et al., 2003 |
|
B |
F5 |
14* (7-22) |
1994-1998 |
(1,4) (0,11-3,4) |
(0,30) (0,05-1,8) |
(0,13) (0,01-1,1) |
(0,14) (<0,01-0,97) |
(4,2) (2,6-6,1) |
Idem |
|
C |
M44 |
42 (22-74) |
1988-2002 |
4,6±1,6 (4,3) (0,78-9,5) |
0,06±0,04 (0,05) (0,02-0,27) |
0,69±0,38 (0,60) (0,13-2,1) |
1,0±0,66 (0,74) (0,04-3,0) |
117±50 (109) (<0,01-260) |
LEBEUF et al., 2007a |
|
C |
F42 |
40 (20-64) |
1987-2002 |
1,3±0,76 (1,1) (0,32-3,2) |
0,08±0,05 (0,06) (0,01-0,22) |
0,20±0,27 (0,13) (0,03-1,5) |
0,71±0,52 (0,50) (0,01-2,4) |
20±18 (15) (<0,01-90) |
Idem |
Foie |
C |
?1 |
<1 |
1991 |
1,5 |
0,24 |
0,67 |
0,10 |
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
2,2 |
0,22 (0,21-0,24) |
2,0 (0,42-4,0) |
0,81 |
|
METCALFE et al., 1999 |
|
C |
M8 |
28 (16-48) |
1995-1999 |
|
|
0,62 (0,19-1,3) |
1,2 (0,08-2,5) |
|
DEVIGNE 2003 |
|
C |
F10 |
38 (10-62) |
1995-1999 |
|
|
0,58 (0,05-3,6) |
0,66 (0,05-2,4) |
|
DEVIGNE 2003 |
|
C |
M3/ F3 |
40 (11-55)/ 5,3 (<1-9) |
2000-2003 |
2,6 ±0,61 (0,06-4,2) |
0,07±0,02 (<0,01-0,14) |
0,12±0,02 (0,02-0,17) |
0,05±0,01 (<0,01-0,08) |
|
McKINNEY et al., 2006 |
Rein |
C |
?1 |
<1 |
1991 |
0,81 |
0,18 |
0,44 |
0,06 |
|
GAUTHIER et al., 1998 |
Cerveau |
C |
?1 |
<1 |
1991 |
0,20 |
0,21 |
0,16 |
0,03 |
|
GAUTHIER et al., 1998 |
|
C |
M4 |
35(3) (28-40) |
1988-1989 |
1,2 |
0,42 (0,29-0,55) |
1,4 (0,28-3,4) |
0,57 |
|
METCALFE et al., 1999 |
Muscle |
C |
M4 |
35(3) (28-40) |
1988-1989 |
4,7 |
0,18 (0,09-0,27) |
2,6 (1,0-6,2) |
1,1 |
|
METCALFE et al., 1999 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal
Tableau 5
Concentrations des polychlorodibenzo-p-dioxines et furannes (PCDD/F), non-ortho et mono-ortho BPC (moyenne ± écart-type, gamme) dans les tissus des bélugas du Saint-Laurent.
Concentrations of polychlorinated dibenzo-p-dioxins and furans (PCDD/F), non-ortho and mono-ortho PCB (mean ± std dev, range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (gamme) |
|
|||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
années |
TCDD/F (pg•g‑1 lip) |
PCDD (pg•g‑1 lip) |
TEQ (PCDD/F) (pg•g‑1 lip) |
Non-ortho PCB (ng•g‑1 lip) |
Mono-ortho PCB (µg•g‑1 lip) |
TEQ (PCB) (pg•g‑1 lip) |
Référence |
Gras |
C |
M5 |
37 (<1-62) |
1988-1990 |
1,4±0,2 (1,1-2,0) |
|
0,14±0,02 (0,11-0,17) |
13,1±9,0 (6,1-27,1) |
7,2±6,6 (1,8-18,2) |
1660±1300 (748-3840) |
MUIR et al., 1996a |
|
C |
F5 |
37 (5-62) |
1987-1990 |
3,0±1,0 (2,1-4,5) |
|
0,30±0,10 (0,21-0,45) |
3,2±2,4 (1,7-7,2) |
1,6±0,67 (0,62-2,2) |
380±220 (135-724) |
idem |
|
C |
?1 |
<1 |
1991 |
|
18 |
0,018 |
0,51 |
1,73 |
255 |
GAUTHIER et al., 1998 |
Foie |
C |
?1 |
<1 |
1991 |
|
1170 |
1,43 |
|
1,0 |
127 |
GAUTHIER et al., 1998 |
Rein |
C |
?1 |
<1 |
1991 |
|
261 |
0,54 |
0,17 |
0,34 |
52 |
GAUTHIER et al., 1998 |
Cerveau |
C |
?1 |
<1 |
1991 |
|
12 |
0,012 |
0,01 |
0,18 |
24 |
GAUTHIER et al., 1998 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
somme des 2,3,7,8-TCDD et 2,3,7,8-TCDF,
somme des 7 PCDD,
concentration en équivalent toxique de la 2,3,7,8-TCDD calculée en multipliant la concentration des congénères détectés par leur facteur d’équivalent toxique respectif (TEF) déterminé par NATO (1988) pour les PCDD/F et AHLBORG et al.(1994) pour les BPC,
somme des congénères non-ortho 77, 126 et 169,
somme des congénères mono-ortho 195, 118 et 156.
Tableau 6
Concentrations des polychloro-n-alkanes (PCA), dieldrine, endrine et octachlorostyrène (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas de l’estuaire du Saint-Laurent.
Concentrations of polychlorinated‑n-alkanes (PCA), dieldrin, endrin et octachlorostyrene (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (moyenne géométrique et/ou gamme) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
PCA (Courte C) (µg•g‑1 lip) |
PCA (Moyenne C) (µg•g‑1 lip) |
Dieldrine (µg•g‑1 lip) |
Endrine (µg•g‑1 lip) |
Octachlorostyrène (ng•g‑1 lip) |
Référence |
Gras |
C |
M4 |
35 (8-47) |
1986-1987 |
|
|
0,93±0,12 (0,81-1,1) |
|
|
MUIR et al., 1990 |
|
C |
F5 |
31 (5-58) |
1986-1987 |
|
|
(0,56±0,31) (0,21-0,87) |
|
|
Idem |
|
C |
?3 |
nd |
1952- 1986 |
|
|
0,50±0,70 (0,05-1,3) |
|
|
JARMAN et al., 1992 |
|
C |
M15 |
37 (<1-62) |
1988- 1990 |
|
|
(1,7) (0,42-5,3) |
|
(36) (<1-186) |
MUIR et al., 1996a |
|
C |
F21 |
37 (5-62) |
1987-1990 |
|
|
(0,63) (0,23-2,9) |
|
(63) (16-179) |
Idem |
|
C |
M9 |
39 (8-56) |
1993-1994 |
|
|
(2,0) (1,4-2,6) |
|
(53) (32-135) |
MUIR et al., 1996b |
|
C |
F7 |
39 (20-66) |
1993-1994 |
|
|
(1,4) (0,33-3,3) |
|
(39) (29-50) |
Idem |
|
C |
M10 |
35 (7-45) |
1986-1991 |
55±24 (32-100) |
47±26 (22-93) |
|
|
|
BENNIE et al., 2000 |
|
C |
F15 |
41 (3-62) |
1987-1991 |
30±22 (5-71) |
24±26 (2-93) |
|
|
|
Idem |
|
C |
M3 |
45 (40-54) |
1988-1989 |
1,0±0,55 (0,49-1,6) |
|
|
|
|
TOMY et al., 2000 |
|
C |
F2 |
54 (46-62) |
1988 |
0,84 (0,69-1,0) |
|
|
|
|
Idem |
|
B |
M34 |
14* (8-22) |
1994-1998 |
|
|
(0,60) (0,04-8,5) |
(0,26) (0,04-2,0) |
(48) (<1-508) |
HOBBS et al., 2003 |
|
B |
F10 |
14* (7-22) |
1994-1998 |
|
|
(0,20) (0,02-1,1) |
(0,16) (<0,01-0,86) |
(24) (<1-623) |
Idem |
Foie |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
|
1,1 |
0,05 |
|
METCALFE et al., 1999 |
|
C |
M3 |
27 (4-40) |
1988 |
22±8,7 (12-29) |
14±2,8 (11-17) |
|
|
|
BENNIE et al., 2000 |
|
C |
F3 |
11 (<1-32) |
1988 |
52±66 (4,9-128) |
30±35 (5,0-70) |
|
|
|
Idem |
|
C |
M3/ F3 |
40 (11-55)/ 5.3 (<1-9) |
2000-2003 |
|
|
0,76±0,19 (0,03-1,2) |
|
11±4 (<0,5-23) |
McKINNEY et al., 2006 |
Cerveau |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
|
0,55 |
0,01 |
|
METCALFE et al., 1998 |
Muscle |
C |
M4 |
35(3) (28-40) |
1988-1989 |
|
|
2,0 |
0,01 |
|
METCALFE et al., 1998 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
âge estimé à partir d’observations répétées des animaux et lorsque possible en référence au changement de couleur de l’animal.
Tableau 7
Concentrations des polybromodiphényles éthers (PBDE) (moyenne ± écart-type, moyenne géométrique et/ou gamme) dans les tissus des bélugas du Saint-Laurent.
Concentrations of polybrominated diphenylethers (PBDE) (mean ± std dev, geometric mean and/or range) in tissues of the St. Lawrence belugas.
|
|
|
|
|
Moyenne ± écart-type (gamme) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|
Tissu |
État |
Sexe, N |
CdC moyenne (gamme) |
Années |
BDE-47 (µg•g‑1 lip) |
BDE-99 (µg•g‑1 lip) |
BDE-100 (µg•g‑1 lip) |
Σ5PBDE (µg•g‑1 lip) |
ΣPBDE (µg•g‑1 lip) |
Référence |
Gras |
C |
M7 |
29 (18-38) |
1999-2000 |
0,28 |
0,11 |
0,06 |
|
0,54 |
LAW et al., 2003 |
|
C |
F8 |
30 (17-54) |
1999-2000 |
0,26 |
0,10 |
0,06 |
|
0,54 |
Idem |
|
C |
M28 |
39 (18-58) |
1988-1999 |
0,15±0,11 (0,01-0,39) |
0,05±0,05 (<0,01-0,20) |
0,03±0,03 (<0,01-0,09) |
0,28±0,20 (0,03-0,73) |
0,29±0,21 (0,03-0,78) |
LEBEUF et al., 2004 |
|
C |
F26 |
42 (20-63) |
1988-1999 |
0,17±0,15 (0,01-0,54) |
0,07±0,06 (<0,01-0,22) |
0,04±0,04 (<0,01-0,13) |
0,33±0,28 (0,02-1,0) |
0,34±0,29 (0,02-1,1) |
Idem |
Foie |
C |
M3/ F3 |
40 (11-55)/ 5,3 (<1-9) |
2000-2003 |
|
|
|
2,0±0,43 (0,21-2,8) |
2,20±0,66 (0,25-3,0) |
McKINNEY et al., 2006 |
carcasse,
biopsie sur animal vivant,
nombre,
mâle,
femelle,
couches de croissance,
congénères BDE‑47, ‑99, ‑100, ‑153 et ‑154