
Résumés
Résumé
En complément aux travaux réalisés depuis 1994 sur la toxicologie des cyanobactéries dans différents lacs-réservoirs du Maroc, le présent travail se propose d'apporter des informations supplémentaires en se fixant trois principaux objectifs: 1) Mise à jour de la base de données de la variation temporelle des teneurs en cyanotoxines intracellulaires (microcystines) d'efflorescences cyanobactériennes fréquemment occasionnés dans le lac réservoir Lalla Takerkoust; 2) Évaluation de la contamination des eaux brutes du lac par les microcystines (MC) (quantification des MC extracellulaires); 3) Caractérisation de la diversité moléculaire des souches cyanobactériennes par la détection des gènes de la synthèse des cyanotoxines (MC) en utilisant les deux méthodes multiplex‑PCR et RFLP (Restriction Fragment Length Polymorphism). L'analyse par HPLC des échantillons 2005 et 2006 d'efflorescences cyanobactériennes a montré qu'il y a une variation qualitative et quantitative des microcystines intracellulaires (MC). L'évaluation des teneurs en MC et dissoutes dans l'eau brute, par ELISA, a révélé des quantités très importantes de MC extracellulaires avec un maximum de 95,4 μg•L‑1 durant le mois de décembre 2005 (phase de déclin du développement des cyanobactéries). En général, durant l'année, les concentrations des MC dissoutes restent toujours au‑dessus de la valeur guide recommandée par l'OMS pour l'eau de boisson (1 μg•L‑1). La caractérisation moléculaire, recherche du gène de synthèse des MC, a confirmé que seule Microcystis aeruginosa est la souche productrice des MC au sein de la fraction phytoplanctonique. Ce travail a pour mérite de confirmer pour la première fois au Maroc que, lors de proliférations d'efflorescences cyanobactériennes à Microcystis, les teneurs en MC dans les eaux brutes du lac sont si importantes qu'il est fortement recommandé de prendre en compte les divers risques sanitaires potentiellement engendrés par cette contamination lors de l'utilisation de cette eau (eau d'alimentation sans traitement, eau récréative, eau d'irrigation, etc.). D'après ces résultats, nous concluons que la mise en place d'un programme de surveillance des cyanobactéries et de contrôle des cyanotoxines (MC) doit être basée sur la caractérisation biochimique (détermination de la nature et de la quantité de toxines produites) et complémentée par la caractérisation génétique des souches potentiellement productrices de ces cyanotoxines.
Mots-clés :
- Microcystis aeruginosa,
- microcystines,
- contamination,
- mcyA,
- multiplex-PCR,
- RFLP
Abstract
Complementary to earlier research carried out since 1994 related to toxic cyanobacteria in different Moroccan lake-reservoirs, this work furnishes new data relating to this research topic. Three major objectives were established: 1) update knowledge concerning the temporal variation of the intracellular contents of cyanotoxines (microcystines, MCs) in cyanobacteria blooms, which frequently occur in the Lalla Takerkoust lake-reservoir; 2) evaluate the MC contamination of raw waters (quantification of extracellular MCs); 3) molecular characterization of cyanobacteria MC producing strains (detection of genes involved in MC synthesis using two methods, multiplex-PCR and RFLP (Restriction fragment length polymorphism)). For bloom samples collected during 2005-2006, the HPLC analysis showed quantitative and qualitative temporal variation in intracellular MCs. Evaluation of the extracellular MC content, determined by ELISA analysis, revealed a maximum concentration of 95.4 μg•L-1 in raw water. This value was measured during December 2005, corresponding to the decline of a Microcystis bloom (cell lysis and MC release into the surrounding water). In general, the dissolved MC concentrations in raw water always remained above the guideline value recommended by the World Health Organization for drinking water (1 μg•L-1). The genetic analysis, based on the detection of the MC synthetase gene ( mcyA gene), confirmed that among the bloom-forming phytoplanktonic fraction, Microcystis aeruginosa is the only MC producing strain. This work thus confirms for the first time in Morocco that during Microcystis blooms, the MC extracellular content greatly exceeds WHO guidelines. It is strongly recommended that the various potential sanitary risks involved in the use of such untreated water (drinking, recreation and irrigation waters) be taken into account. Based on these results, we conclude that the establishment of any successful monitoring programs for cyanobacteria and MC should be based first on biochemical characterization (identification and determination of the amount of produced toxins) and be supplemented by the genetic characterization of the toxin-producing strains.
Keywords:
- Microcystis aeruginosa,
- microcystins,
- contamination,
- mcyA,
- multiplex-PCR,
- RFLP
Corps de l’article
Introduction
La prolifération massive des cyanobactéries est souvent accompagnée de la libération de certaines substances toxiques, métabolites secondaires appelés « cyanotoxines ». Ces biotoxines sont à l’origine de nombreuses nuisances écologiques, sanitaires et socio-économiques qui se manifestent en ce qui a trait à l’altération de la qualité, à la réduction de la productivité et à la limitation d’usage des milieux aquatiques (WIEGAND et PFLUGMACHER, 2005). Au Maroc, la prolifération des cyanobactéries a été largement suspectée dans différents plans d’eau, cependant, les études sur leur toxicologie et leur implication écologique et sanitaire sont encore très rares (OUDRA et al., 2002a, 2002b). Le lac Lalla Takerkoust (bassin versant de N’fis, Marrakech) constitue le premier hydrosystème marocain où des études écotoxicologiques des efflorescences à cyanobactéries ont été réalisées (OUDRA et al., 1998, 2002a; SBIYYAA et al., 1998). Vu l’intérêt écologique et socio-économique que ce lac-réservoir représente dans la région aride et semi-aride de Marrakech-Tensift-Haouz, ce présent travail a le mérite de compléter et faire une mise à jour de la base de données sur l’évolution de ce phénomène de prolifération massive des cyanobactéries et fournir des informations toxicologiques et toxicologiques récentes et utiles pour la mise en place d’un programme moderne de surveillance des cyanobactéries et de contrôle des cyanotoxines, basé sur l’application de nouvelles techniques biologiques nécessaires pour un diagnostique fiable. Ce travail présente donc l’originalité de faire la caractérisation biochimique des toxines intracellulaires et extracellulaires (évaluation de la contamination des eaux brutes du lac par les cyanotoxines de type microcystines (MC), information nécessaire pour les gestionnaires des usines de production de l’eau potable). En outre, cette partie est complémentée par la caractérisation génétique des souches potentiellement productrices de ces cyanotoxines (MC) (contrôle basé sur la détection des gènes de biosynthèse des microcystines, outil moléculaire susceptible d’apporter des réponses rapides sur la toxicité potentielle du phytoplancton).
2. Matériel et méthodes
2.1 Description du site d’étude
Le barrage Lalla Takerkoust est situé à environ 35 km au sud-ouest de la ville de Marrakech (31°36’N, 8°2’W). C’est un lac eutrophe à hypereutrophe, peu profond et sans stratification thermique stable (MOUHRI et al., 1999). Comme la plupart des retenues de barrages eutrophes du Maroc, le lac-réservoir Lalla Takerkoust connaît fréquemment, et quasi annuellement, des proliférations massives de cyanobactéries en été et en début d’automne qui se caractérisent par la formation d’efflorescences cyanobactériennes ou écumes en surface de l’eau et qui, sous l’effet du vent dominant, se retrouvent accumulées au niveau des berges de la retenue. (OUDRA, 2001).
2.2 Échantillonnage et collecte d’efflorescences cyanobactériennes (2003-2006)
Au niveau des berges, la biomasse cyanobactérienne est concentrée à l’aide d’un filet à plancton de 27 µm de vide de maille. La biomasse récoltée est partiellement congelée à -25 °C; l’autre partie est lyophilisée en vue d’extraire et prépurifier les cyanotoxines hépatotoxiques (microcystines).
2.3 Mise en culture de la souche M. aeruginosa
En vue de faire le criblage des souches cyanobactériennes responsables de la formation d’efflorescences cyanobactériennes, après récolte du phytoplancton au filet, un premier inoculum est mis en culture dans des boîtes de pétri contenant le milieu de culture solide Z8 (KOTAI, 1972). Après plusieurs repiquages, la souche dominante de M. aeruginosa est parfaitement isolée et puis cultivée en biomasse. Cette culture monospécifique est réalisée dans des flacons de 500 mL sous des conditions bien contrôlées de luminosité (82 µE•m-2•s-1) et de température (25 ± 2 °C). La souche est maintenue en culture sous aération continue avec une photopériode de 16/8 h. Après cinq jours de culture, la biomasse est récoltée, lyophilisée et ensuite conservée à -25 °C; elle servira pour extraire les toxines. Par contre, la culture mère est maintenue en phase exponentielle de croissance par des repiquages successifs tous les deux à trois jours.
2.4. Caractérisation des microcystines dans les échantillons d’efflorescences prélevés
2.4.1 Extraction et prépurification des microcystines
Les microcystines (MC) sont extraites et prépurifiées selon la méthode préalablement décrite par LAWTON et al. (1994). En bref, une quantité de 250 mg de lyophilisat d’efflorescence cyanobactérienne naturelle ou d’isolat de Microcystis a été extraite trois fois par le méthanol 70 %. Après chaque extraction, la suspension est centrifugée à 4 000 xg (10 min, 4 °C). L’extrait total est dilué avec de l’eau ultrapure (Milli-Q, Millipore) de façon à obtenir un extrait à 20 % de méthanol. Pour la prépurification des MC, l’extrait final est passé dans une colonne de gel de silice ODS (Sep-pak Vac C18, Waters Corporation, Milford, MA, USA). La dernière fraction récupérée contenant les MC est complètement évaporée à 40 °C et mise en solution dans 1 mL de méthanol/eau ultra-pure (50:50, v/v) et filtrée sur un filtre de 0,2 µm (Acrodisc, Nylon, Gelman Sciences Inc.) avant d’être analysée par HPLC.
2.4.2 Quantification et identification des MC par HPLC-PDA
L’analyse des MC contenues dans les extraits de la biomasse cyanobactérienne a été réalisée à l’aide de la Chromatographie Liquide Haute Performance (HPLC, Waters Corporation, modèle 2695) avec un détecteur à barrette photodiode (modèle 996). Les variantes de MC sont identifiées sur la base de leurs temps de retention et leurs spectres d’absorption UV (LAWTON et al., 1994). La colonne utilisée est une Chromolith C18 (250 mm x 4,6 mm, 5 μm) (Waters, Chromatography Division/Millipore Corp.). La phase mobile du système est (A) : H2O + l’acide trifluoroacétique (TFA) 0,05 % (v/v), et (B) : Acetonitrile (MeCN) + TFA 0,05 % (v/v). Le volume de l’échantillon injecté est de 50 μL à 100 μL. La phase mobile fonctionne à 1 mL•min-1. Les standards de MC-LR, -YR, -RR, -LF et -LW sont commercialisés par Calbiochem (Allemagne); quant aux standards de MC-FR et -WR, ils sont purifiés au Laboratoire de physiologie végétale de l’Université Autonome de Madrid. Les résultats de la quantification des MC sont exprimés en équivalents MC-LR en ajoutant la masse de toutes les variantes détectées dans l’extrait. Tous les produits chimiques utilisés sont de qualité chromatographique (Scharlau Chimie Barcelone, Espagne).
2.5 Détermination de la concentration des MC dans l’eau brute
2.5.1 Prélèvement et préparation des échantillons d’eau brute
Afin de mettre en évidence la contamination de l’eau du lac-réservoir Lalla Takerkoust par les MC, un prélèvement mensuel de l’eau brute a été effectué durant l’année 2006 au niveau d’un seul point situé loin de la zone de formation d’efflorescences cyanobactériennes. Un volume de 200 mL de l’échantillon d’eau est filtré sous vide sur un filtre GF/C. Le filtrat est ensuite passé à travers une colonne d’ODS-C18 (Sep-pak Vac C18, Waters Corporation). Les toxines sont récupérées après élution par 20 mL du méthanol 100 %, puis la fraction est évaporée à 40 °C. Le résidu sec est resuspendu dans 1 mL de méthanol.
Le filtre, sur lequel est retenue la fraction sestonique, est extrait avec 20 mL de méthanol 100 % puis sonifié pendant cinq minutes afin de lyser les cellules et libérer les endotoxines. L’extrait est ensuite refiltré sur filtre GF/C et évaporé à sec à 40 °C. Le résidu sec est repris dans 1 mL de méthanol (MAATOUK et al., 2002).
2.5.2 Quantification des MC totales dans l’eau par ELISA
La quantification des MC totales contenues dans l’eau brute filtrée et concentrée a été réalisée par le test immunologique ELISA (Envirogard® Microcystins Plate Kit, Strategic Diagnostic, Newark, DE, USA) selon la procédure décrite par CHU et al. (1990). L’absorbance est déterminée sur un lecteur ELISA DENLEY We-Scan à 450 nm. Les résultats sont reportés en équivalents microcystine-LR. (µg Equiv. MC-LR•L-1).
2.6 Caractérisation génétique d’efflorescences cyanobactériennes par multiplex-PCR et RFLP
2.6.1 Analyse multiplex-PCR
Pour chaque échantillon, une petite quantité d’efflorescences cyanobactériennes est placée dans un tube à centrifugation de 1,5 mL. L’ADN est extrait avec le Kit AquaPure Genomic DNA Isolation (Bio-Rad, Hercules, CA, USA) en suivant les instructions données pour les bactéries gram-négatif. La concentration et la pureté de l’extrait d’ADN sont confirmées par spectrophotométrie à 260 et 280 nm. Les extraits d’ADN sont ensuite testés en utilisant les amorces décrites dans le tableau 1. La paire d’amorces Micr184F/Micr431R est utilisée pour amplifier le fragment spécifique à Microcystis (220-pb) du gène ARNr16s, confirmant la présence de Microcystis dans l’échantillon analysé (NEILAN et al., 1997). La paire d’amorces mcyA-Cd1F/ mcyA-Cd1R est utilisée pour amplifier le fragment du gène mcyA (297-pb) présent dans les espèces productrices des MC et appartenant aux genres Anabaena, Microcystis et Planktothrix (HISBERGUES et al., 2003). Les échantillons ayant donné des résultats positifs dans cette analyse sont, fort probablement, des producteurs de MC (HISBERGUES et al., 2003; SAKER et al., 2005b, 2006). Ces deux paires d’amorces diffèrent par leur température optimale d’hybridation (de 50 à 60 °C). Des expériences préliminaires ont confirmé que 56 °C est la température appropriée pour maximiser le rendement et la spécificité des produits. Le programme AutoDimer (VALLONE et BUTLER, 2004) a été employé pour déterminer l’activité hétérospécifique des quatre amorces incluses dans l’analyse multiplex.
Tableau 1
Amorces utilisées dans l’analyse multiple-PCR
Primers used in the multiplex-PCR analysis

Toutes les réactions de PCR ont été effectuées dans un volume de 20 μL contenant 1 × tampon PCR, 2,5 mM MgCl2, 250 μM de chaque désoxynucléotide triphosphate, 10 pmol de chaque amorce, 0,5 U de Taq ADN polymérase et 5-10 ng d’ADN. Le cycle thermique est effectué dans un Bio-Rad MyCycler (Bio-Rad, Hercules, CA, USA) avec une dénaturation initiale à 95 °C durant 2 min suivie de 35 cycles de 95 °C durant 90 s, 56 °C durant 30 s et 72 °C durant 50 s, et une étape finale supplémentaire à 72 °C pendant 7 min. Quatre microlitres du mélange de la réaction PCR sont déposés pour migration sur gel d’agarose 1,5 % dans 1 × tampon TAE. Les gels ont été souillés avec du bromure d’éthidium et photographiés sous transillumination UV (SAKER et al., 2007).
2.6.1 Analyse RFLP
Afin de déterminer la souche responsable de la production des MC dans les échantillons d’efflorescences cyanobactériennes, une analyse RFLP (Restriction Fragment Length Polymorphism) est appliquée sur les produits de PCR résultant de la paire d’amorces mcyA-Cd1F/mcyA-Cd1R. Premièrement, 18 µL des produits de PCR sont purifiés pour éliminer les désoxynucléotides triphosphates non incorporés et les autres produits réactionnels par combinaison avec 36 µL d’éthanol 95 % et 1,8 µL d’acétate de sodium 3 M. La solution est ensuite vortexée et centrifugée à 13 200 rpm pendant 20 min. Les produits de PCR sont ensuite resuspendus dans 18 µL d’eau ultrapure stérile et digerés avec l’EcoRV à 37 °C. Les produits de PCR digerés sont analysés avec du gel d’agarose 2 % en utilisant le protocole d’électrophorèse standard (SAKER et al., 2007).
3. Résultats et discussion
3.1 Caractérisation des MC dans les échantillons d’efflorescences cyanobactériennes naturelles et dans la culture de M. aeruginosa réalisée au laboratoire
La quantification et la caractérisation par HPLC des extraits d’efflorescences cyanobactériennes ont montré la présence de MC en quantités importantes (Tableau 2). L’analyse montre qu’il y a une variation temporelle, aussi bien quantitative que qualitative, des MC dans les échantillons récoltés du lac-réservoir Lalla Takerkoust de 2005 à 2006 (Tableau 2; Figure 1). La concentration de MC la plus élevée est enregistrée en décembre 2005, avec une valeur de 3 240 µg•g-1 du poids sec (ps). Dans la même année, cette concentration est de 1 160 et 519 µg•g-1 ps respectivement en août et septembre. En juillet 2006, cette concentration atteint 759 µg•g-1 ps. Les chromatogrammes des extraits analysés montrent la détection de plusieurs variantes (MC-RR, -(H4)YR, -YR, -LR, -FR et -WR) dont le pourcentage de dominance varie en fonction du temps (Figure 1). Parmi ces variantes, on trouve que la MC-RR (33 à 35 %) et MC-LR (24 à 32 %) sont les plus dominantes, tandis que la MC-FR (2 à 3 %) et MC-WR (2 à 4 %) sont les moins répandues. L’analyse de l’extrait d’isolat de Microcystis révèle que la culture au laboratoire n’a pu produire que deux variantes de MC qui sont la MC-RR (85,1 %) et MC-WR (14,9 %) (Tableau 2; Figure 2). En comparaison avec les résultats de l’analyse d’efflorescences cyanobactériennes, la teneur en MC totales dans l’extrait de l’isolat de Microcystis est 40 fois plus faible que celle de l’efflorescence cyanobactérienne prélevée en décembre 2005.
Pour certains lacs-réservoirs d’Afrique du Sud, WICKS et THIEL (1990) ont rapporté des valeurs comprises entre 5 et 630 µg•g-1. Dans des lacs-réservoirs au Portugal, ces valeurs sont comprises entre 1 000 et 1 700 µg•g-1 (VASCONCELOS, 1993). En France, ces valeurs varient de 30 à 3 970 µg•g-1 (VEZIE et al., 1997, 1998). Ces teneurs montrent clairement la forte variation des toxines en fonction de la situation géographique et des conditions environnementales. En effet, il y a quelques facteurs qui ont une influence directe sur la synthèse des MC. Généralement, cette synthèse augmente avec l’intensité lumineuse (KIM et al., 2005) mais la qualité de la lumière est également un facteur déterminant (la lumière rouge favorise la production de toxines alors que la lumière bleue ne le fait pas) (KAEBERNICK et al., 2000). Il a été montré aussi que la température influence le type de toxines produites, avec les températures élevées (>25 °C) augmentant la production de MC-RR et les plus basses températures favorisant la synthèse de MC-LR (RAPALA et al., 1997; RAPALA et SIVONEN, 1998). En plus, la production de MC par les souches de Microcystis semble être influencée par la variation des concentrations en azote et en phosphore avec différentes réponses selon la souche considérée (VEZIE et al., 2002).
tableau 2
Résultats de l’analyse HPLC-PDA
HPLC-PDA analysis results

Figure 1
Chromatogrammes de l’analyse HPLC-PDA effectuée sur des extraits d’efflorescences cyanobactériennes M. aeruginosa, collectés du lac-réservoir Lalla Takerkoust en 08/2005 (A), 09/2005 (B), 12/2005 (C) et 07/2006 (D) avec leurs spectres UV
Chromatograms of the HPLC-PDA analysis carried out on M. aeruginosa bloom extracts collected from Lalla Takerkoust lake-reservoir in 08/2005 (A), 09/2005 (B) 12/2005 (C) and 07/2006 (D) with their UV spectra

Figure 2
Chromatogramme de l’analyse HPLC-PDA effectuée sur l’extrait de la culture de M. aeruginosa. La souche à été isolée du lac-réservoir Lalla Takerkoust en 2003 et cultivée au laboratoire
Chromatogram of the HPLC-PDA analysis carried out on the M. aeruginosa culture extract. The strain was isolated from the Lalla Takerkoust in 2003 and cultivated at the laboratory
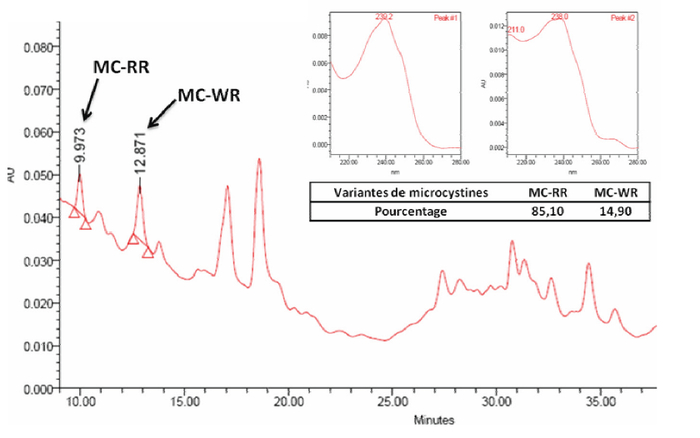
Figure 3
Suivi des concentrations de microcystines dans l’eau brute du lac-réservoir Lalla Takerkoust durant l’année 2006
Monitoring of microcystin concentrations in raw water of the Lalla Takerkoust lake-reservoir during the year 2006

3.2 Suivi de la concentration de MC dans l’eau du lac de barrage Lalla Takerkoust
L’analyse par ELISA a montré la présence de MC aussi bien en période de prolifération d’efflorescences cyanobactériennes (été-automne) que pendant les mois marquant l’absence totale de prolifération, avec une concentration moyenne de 28,9 µg•L-1. La figure 3 montre que les teneurs en MC sont variables en fonction des saisons. Les valeurs les plus fortes sont enregistrées pendant les saisons estivale et automnale. Ainsi, des pics remarquables ont été enregistrés pendant les mois de décembre de l’année 2005 et en septembre de l’année 2006 respectivement à des concentrations d’environ 95,4 et 60 µg•L-1. Ces concentrations importantes sont dues à la lyse des cellules de cyanobactéries après une prolifération intense. En effet, les MC sont censées être maintenues principalement dans les cellules des cyanobactéries et ne peuvent être libérées dans l’eau qu’après la sénescence et la décomposition des cellules (WIEGAND et PFLUGMACHER 2005).
C’est la première fois qu’un suivi des concentrations de MC dans l’eau brute d’un lac de barrage est réalisé au Maroc. Durant toute l’année 2006, la concentration de MC dans l’eau reste toujours au-dessus de la barre de 1 µg•L-1 qui est fixée par l’OMS comme concentration maximale réglementaire dans l’eau de boisson (OMS, 2004).
3.3 Caractérisation génétique d’efflorescences cyanobactériennes par multiplex-PCR et RFLP
La figure 4 montre les résultats obtenus à partir de l’analyse PCR réalisée sur des échantillons d’efflorescences cyanobactériennes naturelles prélevés dans le lac de barrage Lalla Takerkoust en 2005 et 2006. Une synthèse des résultats de cette analyse figure dans le tableau 3. Tous les échantillons testés ont donné des résultats positifs pour la présence de Microcystis indiquée par la formation de produits de PCR à 220-pb (Figure 4). Les deux paires d’amorces Cyl2/Cyl4 et PKS M4/PKS M5 utilisées dans l’analyse PCR ont toutes les deux donné des résultats négatifs avec les extraits d’ADN de tous les échantillons d’efflorescences cyanobactériennes testés (Figure 5), indiquant l’absence de Cylindrospermopsis et du gène de synthèse des cylindrospermopsines dans tous les échantillons.
Ces résultats sont en total accord avec l’étude microscopique faite sur les mêmes échantillons montrant que Microcystisaeruginosa Kütz est l’espèce dominante dans les échantillons d’efflorescences cyanobactériennes prélevés. Les extraits d’ADN testés avec la paire d’amorces mcyA-Cd1F/mcyA-Cd1R ont tous donné des résultats positifs, avec la production de fragments de gène à 297-pb (Tableau 3, Figure 4). Ce gène est présent chez les souches productrices de MC appartenant aux genres Microcystis, Planktothrix et Anabaena. Une analyse de restriction (RFLP) a donc été appliquée afin de déterminer les espèces responsables de la production des MC dans les échantillons d’efflorescences cyanobactériennes. Après 3 h de digestion à 37 °C, on remarque une différence claire dans les modèles RFLP obtenus pour les souches productrices de MC, Microcystis (M6) et Planktothrix (Cya126) (Figure 6). La ligne M6 montre un modèle RFLP obtenu pour la souche Microcystis pure montrant un seul produit à 220-pb. Par contre, la souche pure de Planktothrix (ligne Cya126) donne un produit d’environs 300-pb. Quand les extraits d’ADN appartenant à Microcystis et Planktothrix sont mélangés ensemble, le modèle de bande résultant après digestion par l’enzyme de restriction montre clairement deux produits de différentes tailles (ligne M6 + Cya126). Dans tous les cas, les profils obtenus dans le test de restriction semble être similaire à celui obtenu avec la souche pure de Microcystis (Figure 6), indiquant que Microcystis est la souche responsable de la synthèse des MC dans tous les échantillons d’efflorescences cyanobactériennes naturelles prélevés dans le lac de barrage Lalla Takerkoust.
Figure 4
Gel d’agarose obtenu à partir da l’analyse multiplex-PCR en utilisant les paires d’amorces Micr184F/Micr431R et mcyA-Cd1F/mcyA-Cd1R. Ligne 1, efflorescence cyanobactérienne 23-08-2005; ligne 2, efflorescence cyanobactérienne 16-09-2005; ligne 3, efflorescence cyanobactérienne 28-09-2005; ligne 4, efflorescence cyanobactérienne 26-12-2005; ligne 5, efflorescence cyanobactérienne 26-07-2006
Agarose gel obtained from multiplex-PCR analysis using the Micr184F/Micr431R and mcyA-Cd1F/mcyA-Cd primer pairs. Line 1, bloom 23-08-2005; line 2, bloom 16-09-2005; line 3, bloom 28-09-2005; line 4, bloom 26-12-2005; line 5, bloom 26-07-2006

Tableau 3
Présence ou absence des gènes cibles dans les extraits d’ADN des échantillons d’efflorescences cyanobactériennes naturelles prélevés dans le lac de barrage Lalla Takerkoust durant les années 2005 et 2006
Presence or absence of target genes in DNA extracts of natural bloom samples collected in the Lalla Takerkoust lake-reservoir between 2005 and 2006

Figure 5
Gel d’agarose obtenu à partir da l’analyse PCR en utilisant les paires d’amorces Cyl2/Cyl4 (A) et PKS M4/PKS M5 (B). Ligne 1, efflorescence cyanobactérienne 23-08-2005; ligne 2, efflorescence cyanobactérienne 16-09-2005; ligne 3, efflorescence cyanobactérienne 28-09-2005; ligne 4, efflorescence cyanobactérienne 26-12-2005; ligne 5, efflorescence cyanobactérienne 26-07-2006; Cylin, culture pure de Cylindrospermopsis
Agarose gel obtained from PCR analysis using the Cyl2/Cyl4 (A) and PKS M4/PKS M5 (B) primer pairs. Line 1, bloom 23-08-2005; line 2, bloom 16-09-2005; line 3, bloom 28-09-2005; line 4, bloom 26-12-2005; line 5, bloom 26-07-2006; Cylin, Cylindrospermopsis pure culture
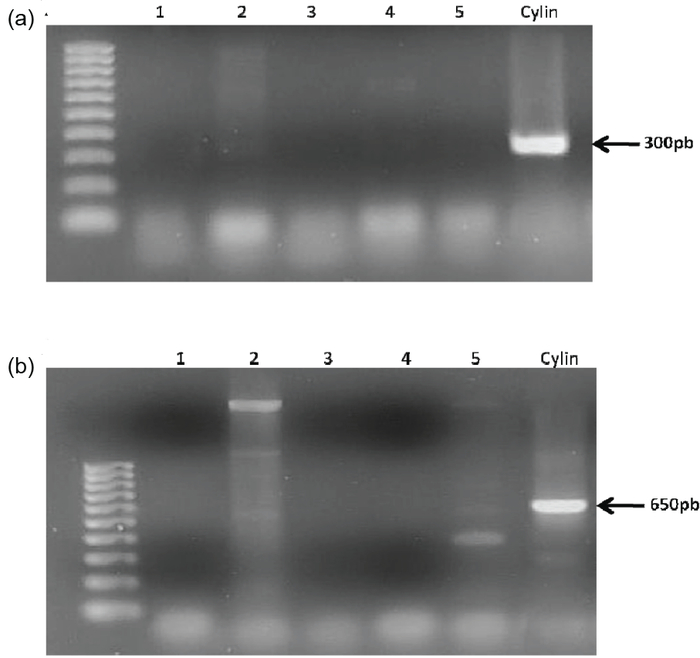
Figure 6
Analyse RFLP des produits de PCR mcyA-CD1 après digestion par l’EcoRV. M6, une souche pure de Microcystis; Cya126, une souche pure de Planktothrix; Ligne 1, efflorescence cyanobactérienne 23-08-2005; ligne 2, efflorescence cyanobactérienne 16-09-2005; ligne 3, efflorescence cyanobactérienne 28-09-2005; ligne 4, efflorescence cyanobactérienne 26-12-2005; ligne 5, efflorescence cyanobactérienne 26-07-2006
RFLP analysis of mcyA-CD1 PCR products digested with EcoRV. M6, Microcystis pure strain; Cya126, Planktothrix pure strain; Line 1, bloom 23-08-2005; line 2, bloom 16-09-2005; line 3, bloom 28-09-2005; line 4, bloom 26-12-2005; line 5, bloom 26-07-2006

4. Conclusion
Le travail a le mérite de faire la combinaison de plusieurs méthodes d’analyses (HPLC-PDA, ELISA, PCR-RFLP) pour le suivi et l’évaluation de la toxicité de l’eau et des efflorescences à cyanobactéries, pour la première fois au Maroc, dans un lac marocain, ce qui constitue une première référence au nord de l’Afrique. L’ensemble des résultats de ce travail montre que les manifestations d’efflorescences dans le lac de barrage Lalla Takerkoust s’accompagnent de la production de MC en quantités relativement élevées. Une partie de ces toxines est libérée dans l’eau avec des concentrations très importantes qui persistent pendant toute l’année. Les analyses moléculaires montrent que Microcystisaeruginosa est la souche responsable de la synthèse de ces toxines dans le lac Lalla Takerkoust, mais il est aussi important de noter que la présence des gènes de la microcystine synthétase dans un échantillon n’implique pas nécessairement la biosynthèse des MC (SAKER et al., 2006). Néanmoins, les méthodes moléculaires (multiplex-PCR et RFLP) utilisées dans ce travail pourraient être employées, dans certains cas, pour remplacer les techniques microscopiques laborieuses et longues. La détection précoce des espèces potentiellement toxiques permettra aux agences responsables de la surveillance de la qualité de l’eau de mettre en place des mesures appropriées pour empêcher la croissance de ces organismes. En plus, des études supplémentaires seront nécessaires pour déterminer les facteurs génétiques et/ou environnementaux régissant la synthèse des MC, surtout au niveau de la variabilité du profil des toxines qui a un impact direct sur la toxicité de la souche cyanobactériale.
Parties annexes
Bibliographie
- CHU F.S., X. HUANG, et R.D. WEI (1990). Enzyme-linked immu-nosorbent assay for microcystins in blue-green algal blooms. J. Assoc. Offic. Anal. Chem., 73, 451-456.
- HISBERGUES M., G. CHRISTIANSEN, L. ROUHIAINEN, K. SIVONEN et T. BÖRNER (2003). PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch. Microbiol., 180, 402-410.
- KAEBERNICK, M., B.A. NEILAN, T. BÖRNER, et E. DITTMAN (2000). Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microbiol., 66, 3387-3392.
- KIM H.R., C.K. KIM, T.S. AHN, S.A. YOO, D.H. LEE (2005). Effects of temperature and light on microcystin synthetase gene transcription in Microcystis aeruginosa. KeyEng. Mater., 277-279, 606-611.
- KOTAI J. (1972). Instruction of preparations of modified nutrient medium Z8 for algae. Norwegian Institue for Water Research, Blindem, Oslo, B-11/69, 5.
- LAWTON L.A., C. EDWARDS et G.A. CODD (1994). Extraction and high-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst., 119, 1525-1530.
- MAATOUK I., N. BOUAÏCHA, D. FONTAN et Y. LEVI (2002). Seasonal variation of microcystin concentrations in the Saint-Caprais reservoir (France) and their removal in a full-scale treatment plant. Water Res., 36, 2891-2897.
- MOUHRI K., M. LOUDIKI, R. LE COHU, N. ZAKI (1999). Effet des enrichissements en nutriments sur le phytoplancton isolé à l’aide d’enceintes expérimentales placées in situ dans deux lacs réservoirs de la région de Marrakech (Maroc). Ann. Limnol., 35, 3-12.
- NEILAN B.A., D. JACOBS, T. DEL DOT, L.L. BLACKALL, P.R. HAWKINS, P.T. COX et A.E. GOODMAN (1997). rRNA sequences and evolutionary relationships among toxic and non-toxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol., 47, 693-697.
- OMS (2004). Guidelines for drinking-water quality. Third edition.Volume 1 - Recommendations. Genève, Suisse. 515 p.
- OUDRA B. (2001). Toxicologie des cyanobactéries (Algues bleu-vert): Cas d’espèces occasionnant des blooms toxiques dans différents milieux aquatiques du Maroc. Thèse de Doctorat d’État, Univ. Cadi Ayyad, Faculté des Sciences-Semlalia Marrakech, Maroc, 118 p.
- OUDRA B., M. LOUDIKI, B. SBIYYAA, B. SABOUR, A. AMORIM, R. MARTINS et V. VASCONCELOS (2002a). Detection and variation of microcystin contents of Microcystis blooms in eutrophic Lalla Takerkoust Lake (Morocco). Lakes Reserv. Res. Manag., 7, 35-44.
- OUDRA B., M. LOUDIKI, B. SBIYYAA, V. VASCONCELOS, H. ZERROUK, M. EL-ANDALOUSSI-DADI et J. DARLEY (1998). Occurrence of hepatotoxic Microcystisaeruginosa blooms in eutrophic Moroccan lake – reservoir. Dans : Harmful Algae, Vigo: Xunta de Galicia. Reguara B., Blanco J., Fernadez M.L., Wyatt T. /IOC-UNESCO (Éditeurs), pp. 29-31.
- OUDRA B., M. LOUDIKI, V. VASCONCELOS, B. SABOUR, B. SBIYYAA, K. OUFDOU et N. MEZRIOUI (2002b). Detection and quantification of microcystins from cyanobacteria strains isolated from reservoirs and ponds in Morocco. Environ. Toxicol., 17, 32-39.
- RAPALA J. et K. SIVONEN (1998). Assessment of environmental conditions that favor hepatotoxic and neurotoxic Anabaena spp. Strains cultured under light limitation at different temperatures. Microb. Ecol., 36, 181-192.
- RAPALA J., K. SIVONEN, C. LYRA et S.I. NIEMELÄ (1997). Variation of microcystins, cyanobacterial hepatotoxins, in Anabaena spp. as a function of growth stimuli. Appl. Environ. Microbiol., 63, 2206-2212.
- SAKER M.L., A-D. JUNGBLUT, B.A. NEILAN, D.F.K. RAWN et V.M. VASCONCELOS (2005). Detection of microcystin synthetase genes in health food supplements containing the freshwater cyanobacterium Aphanizomenon flos-aquae. Toxicon, 46, 555-562.
- SAKER M.L., M. VALE et V.M. VASCONCELOS (2006). Molecular techniques for the early warning of cyanobacteria blooms in freshwater lakes and rivers. Appl. Microbiol. Biotechnol., 75, 441-449.
- SAKER M.L., M. WELKER et V.M. VASCONCELOS (2007). Multiplex PCR for the detection of toxigenic cyanobacteria in dietary supplements produced for human consumption. Appl. Microbiol. Biotechnol., 73, 1136-1142.
- SCHEMBRI M.A., B.A. NEILAN et C.P. SAINT (2001). Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ. Toxicol., 16, 413-421.
- SBIYYAA B., M. LOUDIKI, B. OUDRA, A. BOUGUERNE, A. TIFNOUTI (1998). Acute toxicity of Microcystisaeruginosa kütz. to three cladocerean species. Dans : Harmful Algae. Xunta de Galicia. Reguera B., Blonco J., Fernandez M.L., Wyatt T. IOC-UNESCO (Éditeurs), pp. 382-385.
- VALLONE P.M. et J.M. BUTLER (2004). AutoDimer: a screening tool for primer-dimer and hairpin structures. Biotechniques, 37, 226-231.
- VASCONCELOS V.M. (1993). Toxicity of cyanobacteria in lakes of North and Central Portugal. Ecological implications. Verh. Inter. Verein. Limnol., 25, 694-697.
- VEZIE C, L. BRIENT, K. SIVONEN, G. BERTRU, J.C. LEFEUVRE et M. SALKINOJA-SALONEN (1998). Variation of microcystin content of cyanobacterial blooms and isolated strains in Grand-lieu lake (France). Microb. Ecol., 35, 126-135.
- VEZIE C, L. BRIENT, K. SIVONEN, G. BERTRU, J.C. LEFEUVRE et M. SALKINOJA-SALONEN (1997). Occurrence of microcystin containing cyanobacterial blooms in freshwaters of Brittany (France). Arch. Hydrobiol., 139, 401-413.
- VEZIE C., J. RAPALA, J. VAITOMAA, J. SEITSONEN et K. SIVONEN (2002). Effect of nitrogen and phosphorus on growth of toxic and nontoxic Microcystis strains and on intracellular microcystin concentrations. Microb. Ecol., 43, 443-454.
- WICKS R.J. et P.G. THEIL (1990). Environmental factors affecting the production of peptide toxins in floating scums of the cyanobacterium Microcystis aeruginosa in a hypertrophic African reservoir. Environ. Sci. Technol., 24, 1413-1418.
- WIEGAND C. et S. PFLUGMACHER (2005). Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol. App. Pharmacol., 203, 201-218.
- WILSON K.M., M.A. SCHEMBRI, P.D BAKER et C.P. SAINT (2000). Molecular characterization of the toxic cyanobacterium Cylindrospermopsis raciborskii and design of a species-specific PCR. Appl. Environ. Microbiol., 66, 332–338.
Liste des figures
Figure 1
Chromatogrammes de l’analyse HPLC-PDA effectuée sur des extraits d’efflorescences cyanobactériennes M. aeruginosa, collectés du lac-réservoir Lalla Takerkoust en 08/2005 (A), 09/2005 (B), 12/2005 (C) et 07/2006 (D) avec leurs spectres UV
Chromatograms of the HPLC-PDA analysis carried out on M. aeruginosa bloom extracts collected from Lalla Takerkoust lake-reservoir in 08/2005 (A), 09/2005 (B) 12/2005 (C) and 07/2006 (D) with their UV spectra
Figure 2
Chromatogramme de l’analyse HPLC-PDA effectuée sur l’extrait de la culture de M. aeruginosa. La souche à été isolée du lac-réservoir Lalla Takerkoust en 2003 et cultivée au laboratoire
Chromatogram of the HPLC-PDA analysis carried out on the M. aeruginosa culture extract. The strain was isolated from the Lalla Takerkoust in 2003 and cultivated at the laboratory
Figure 3
Suivi des concentrations de microcystines dans l’eau brute du lac-réservoir Lalla Takerkoust durant l’année 2006
Monitoring of microcystin concentrations in raw water of the Lalla Takerkoust lake-reservoir during the year 2006
Figure 4
Gel d’agarose obtenu à partir da l’analyse multiplex-PCR en utilisant les paires d’amorces Micr184F/Micr431R et mcyA-Cd1F/mcyA-Cd1R. Ligne 1, efflorescence cyanobactérienne 23-08-2005; ligne 2, efflorescence cyanobactérienne 16-09-2005; ligne 3, efflorescence cyanobactérienne 28-09-2005; ligne 4, efflorescence cyanobactérienne 26-12-2005; ligne 5, efflorescence cyanobactérienne 26-07-2006
Agarose gel obtained from multiplex-PCR analysis using the Micr184F/Micr431R and mcyA-Cd1F/mcyA-Cd primer pairs. Line 1, bloom 23-08-2005; line 2, bloom 16-09-2005; line 3, bloom 28-09-2005; line 4, bloom 26-12-2005; line 5, bloom 26-07-2006
Figure 5
Gel d’agarose obtenu à partir da l’analyse PCR en utilisant les paires d’amorces Cyl2/Cyl4 (A) et PKS M4/PKS M5 (B). Ligne 1, efflorescence cyanobactérienne 23-08-2005; ligne 2, efflorescence cyanobactérienne 16-09-2005; ligne 3, efflorescence cyanobactérienne 28-09-2005; ligne 4, efflorescence cyanobactérienne 26-12-2005; ligne 5, efflorescence cyanobactérienne 26-07-2006; Cylin, culture pure de Cylindrospermopsis
Agarose gel obtained from PCR analysis using the Cyl2/Cyl4 (A) and PKS M4/PKS M5 (B) primer pairs. Line 1, bloom 23-08-2005; line 2, bloom 16-09-2005; line 3, bloom 28-09-2005; line 4, bloom 26-12-2005; line 5, bloom 26-07-2006; Cylin, Cylindrospermopsis pure culture
Figure 6
Analyse RFLP des produits de PCR mcyA-CD1 après digestion par l’EcoRV. M6, une souche pure de Microcystis; Cya126, une souche pure de Planktothrix; Ligne 1, efflorescence cyanobactérienne 23-08-2005; ligne 2, efflorescence cyanobactérienne 16-09-2005; ligne 3, efflorescence cyanobactérienne 28-09-2005; ligne 4, efflorescence cyanobactérienne 26-12-2005; ligne 5, efflorescence cyanobactérienne 26-07-2006
RFLP analysis of mcyA-CD1 PCR products digested with EcoRV. M6, Microcystis pure strain; Cya126, Planktothrix pure strain; Line 1, bloom 23-08-2005; line 2, bloom 16-09-2005; line 3, bloom 28-09-2005; line 4, bloom 26-12-2005; line 5, bloom 26-07-2006
Liste des tableaux
Tableau 1
Amorces utilisées dans l’analyse multiple-PCR
Primers used in the multiplex-PCR analysis
tableau 2
Résultats de l’analyse HPLC-PDA
HPLC-PDA analysis results
Tableau 3
Présence ou absence des gènes cibles dans les extraits d’ADN des échantillons d’efflorescences cyanobactériennes naturelles prélevés dans le lac de barrage Lalla Takerkoust durant les années 2005 et 2006
Presence or absence of target genes in DNA extracts of natural bloom samples collected in the Lalla Takerkoust lake-reservoir between 2005 and 2006