

Résumés
Résumé
Contexte L’altération des fonctions émotionnelles ainsi que de la mémoire des personnes atteintes de schizophrénie est connue depuis plus d’un siècle. Toutefois, les structures cérébrales touchées ainsi que les stades de traitement affectés restent encore mal compris.
Objectif Approfondir de quelles façons la valence et l’activation émotionnelle affectent la mémoire épisodique et l’activité cérébrale associée chez des patientes atteintes de schizophrénie.
Hypothèse L’activité des régions frontales associées à certains types de stimuli émotifs sera généralement affectée dans la population clinique, et ce, en réponse aux stimuli à fortes activations émotionnelles.
Participantes Seize patientes atteintes de schizophrénie dans une phase stable de leur maladie furent appariées à 17 participantes contrôles sur la base de l’âge, de la dominance manuelle et du statut socio-économique.
Instrument et mesures L’EEG a été enregistré à partir de 60 électrodes fixées dans un casque en lycra élastique. Ces signaux ont été moyennés pour obtenir les composantes N200, P300, ainsi que la LPC afin de cibler les processus mnésiques et émotionnels.
Résultats Pour la N200 et la P300 frontale, la magnitude de l’effet de mémoire observé chez les patientes schizophrènes est augmentée significativement dans l’hémisphère frontal droit en réponse aux stimuli à valences déplaisantes, alors que l’augmentation de cet effet s’effectue dans les deux hémisphères chez les participantes contrôles.
Conclusion Une altération électrocorticale précoce reflétant la familiarité et la mémoire aux stimuli déplaisants est présente chez les patientes, alors que les processus tardifs de récollection consciente (LPC pariétale) ne seraient pas atteints.
Mots-clés :
- mémoire,
- émotions,
- potentiels évoqués cognitifs,
- schizophrénie,
- ERP
Abstract
Context Abnormal emotion processing is frequent in schizophrenia and affects social and functional outcome. Past event-related potential (ERP) research investigating processing of affective stimuli in schizophrenia was done mainly with facial expressions and revealed impaired facial emotion recognition in patients relative to control subjects. Experimentations involving fMRI with this group of patients, showed alteration of limbic and frontal regions in response to emotional unpleasant images, compared to neutral stimuli during a memory task. Other studies have also noted an increase in brain activity when the activation of the stimuli was high compared to low arousal stimuli. This may indicate a different sensitivity threshold to emotional arousal and emotional valence involving frontal pathways in these patients. But very few studies attempted to separate the contributions of emotional valence and arousal within an episodic memory protocol with ERP, in that population.
Goal The aim of the current research is to investigate brain electro-cortical activity in schizophrenia in response to emotional images during an episodic memory task.
Method ERP components were analyzed in 16 schizophrenic and 17 control participants matched for age, sex and intelligence. ERPs were obtained from 56 EEG electrodes. The tasks consisted in a classical episodic memory task that presented 100 repeated old and 100 new photographic images divided into four categories (unpleasant-high arousal, unpleasant-low arousal, pleasant-high arousal and pleasant-low arousal) selected from the International Affective Picture System. The N200, P300 and late positive component (LPC) mean amplitude, were analyzed using repeated-measure analyses of variance (MANOVA).
Results Patients with schizophrenia and control subjects gave comparable subjective evaluations of arousal and valence. However, the frontal N200 and the P300 both showed an interaction of the group x memory x valence x hemisphere (F [1.32]=8.36; p <.01). Thus, this complex interaction denotes an increase of the episodic memory effect in the right hemisphere in response to unpleasant stimuli, with schizophrenic patients. With respect to the control group, there is also an increase of this memory effect in the right hemisphere, but in response to pleasant stimuli. The schizophrenic patients presented a smaller LPC memory effect, especially at the frontal region. More specifically, the frontal LPC was reduced, and the clinical group was less reactive to the emotional arousal content, compared to the control group.
Discussion Altogether, our results revealed that while the subjective evaluation of emotional pictures is equivalent across groups, cerebral differences are present in schizophrenic patients during emotional recognition. N200 and P300 results in the frontal region suggest impaired selective attention and episodic memory to unpleasant stimuli in patients, while later processes related to conscious recollection (parietal LPC) are not affected with patients affected with schizophrenia.
Conclusion This finding provides further support for the notion of a possible discrepancy between the subjective experience and the physiological expression of emotions in schizophrenia patients. Those results could open the door to new clinical research investigations in psychiatry, particularly in the comprehension of a relationship between frontal cortex vulnerability and episodic memory often present in psychosis.
Keywords:
- schizophrenia,
- psychosis,
- event-related potentials,
- memory,
- emotion,
- valence,
- arousal
Corps de l’article
Schizophrénie, mémoire et émotion
La schizophrénie est considérée comme un trouble neuropsychiatrique à la fois grave, complexe et encore mal compris au plan cognitif, émotionnel et cérébral. Cette complexité est clairement illustrée par l’hétérogénéité de la symptomatologie de ce groupe clinique, allant des hallucinations, des délires, des discours et des comportements désorganisés, jusqu’à un émoussement de l’affect et un état d’anhédonie (Green, 2006 ; Horan, Blanchard, Clark & Green, 2008). Ainsi, les symptômes associés à la schizophrénie sont habituellement divisés en deux catégories distinctes, soit les symptômes positifs et les symptômes négatifs. En général, les symptômes positifs sont des comportements observables, tels que des hallucinations ou une désorganisation manifeste qui apparaissent avec une forte intensité et une fréquence élevée et qui ne sont pas présentes dans la population en général (Fletcher & Frith, 2009). Quant aux symptômes négatifs, ils indiquent les traits associés principalement à des symptômes d’anhédonie (51 %), d’apathie (44 %), d’émoussement affectif (29 %), d’alogie (17 %) et de détérioration de l’attention (34 %) (Häfner et al., 1993). Les premiers symptômes émergent habituellement durant l’adolescence ou tôt dès l’âge adulte, ce qui conduit souvent à un appauvrissement économique et social et à une détresse marquée, tant pour les patients que pour leurs familles (Folsom et al., 2005). Cet état n’entraîne pas que des conséquences sociales, mais se voit aussi par un effet observable sur la sphère cognitive, notamment sur la mémoire épisodique et aussi sur le traitement des émotions qui jouent un rôle capital dans l’expression des symptômes et dans le rétablissement éventuel.
Influences connues des émotions sur la mémoire épisodique
La mémoire épisodique est la mémoire des événements qui s’inscrivent dans un contexte spatial, temporel et émotionnel. Cette mémoire contient ainsi des informations permettant de les replacer dans un contexte particulier (Kim, Vallesi, Picton & Tulving, 2009). Elle permet de répondre à des questions concernant son passé, ses antécédents médicaux, son dernier voyage, son meilleur match de hockey, une scène ou une image perçue, etc. Les résultats d’une méta-analyse, effectuée en imagerie par résonance magnétique fonctionnelle, soulignent que les régions impliquées dans l’encodage sont principalement l’hippocampe gauche antérieur, la partie droite de l’amygdale et le gyrus frontal inférieur (Spaniol et al., 2009), alors que pour la récupération, le gyrus parahippocampique droit, le cortex préfrontal médian (CPFm) et supérieur, le sillon intrapariétal ainsi que le lobe pariétal supérieur et inférieur latéral sont aussi impliqués.
Quant au traitement des émotions en mémoire épisodique, un mécanisme pourrait favoriser la récupération et la consolidation, via une activation amygdalienne, de la formation hippocampique (FH) et du CPFm (Jin & Maren, 2015). Ainsi, la FH et le CPFm interviendraient dans l’encodage et la récupération des souvenirs épisodiques (Preston & Eichenbaum, 2013) et appuieraient ainsi l’hypothèse de l’implication de cette voie dans le traitement de la spécificité contextuelle des souvenirs émotionnels. De plus, cette transmission des signaux contextuels de la FH vers le CPFm générerait une réponse comportementale adaptée au contexte, en interagissant avec le complexe amygdalien (Herry et al., 2008 ; Kalisch et al., 2006 ; Knapska et al., 2012).
Le traitement des émotions et la mémoire dans la schizophrénie
Certaines anomalies entre les structures hippocampiques et frontales ont été signalées chez les patients schizophrènes. Par exemple, ceux-ci exhiberaient un mauvais couplage entre la FH et le CPFm au repos, ainsi que pendant l’exécution d’une tâche de mémoire de travail (Lett, Voineskos, Kennedy, Levine & Daskalakis, 2014 ; Meyer-Lindenberg et al., 2005 ; Zhou et al., 2008). Cela a été confirmé dans des modèles animaux de la schizophrénie, qui présentent un asynchronisme de l’activité de l’EEG quantitatif entre la FH et le CPFm (Sigurdsson, Stark, Karayiorgou, Gogos & Gordon, 2010).
Chez l’humain, les troubles de la mémoire épisodique représentent un déficit important dans la schizophrénie, ce qui limite considérablement le devenir fonctionnel des patients (Reichenberg & Harvey, 2007). À ce titre, deux méta-analyses avec des études en imagerie cérébrale fonctionnelle, contrastant des patients atteints de schizophrénie et des volontaires sains, durant l’encodage et la récupération épisodique (Achim & Lepage, 2005 ; Ragland et al., 2009) ont montré un dysfonctionnement préfrontal proéminent en région dorsolatérale et ventrolatérale, ce qui pourrait suggérer que des déficits de contrôle cognitif contribuent fortement à l’altération de la mémoire épisodique dans la schizophrénie. Toutefois, une mauvaise communication entre les structures hippocampiques et frontales pourrait bien expliquer les déficits en mémoire, mais ceci reste toutefois insuffisant pour expliquer les mécanismes de régulation émotionnelle observés dans la psychose.
L’altération des fonctions émotionnelles chez les personnes atteintes de schizophrénie est connue depuis plus d’un siècle, mais à la fin des années 1990, des études ont confirmé une altération de la sphère émotionnelle chez ces patients, comparativement à d’autres groupes faisant partie de la population psychiatrique, telle que les troubles de l’humeur (Addington & Addington, 1998 ; Gaebel & Wolwer, 1992). Les dimensions touchées incluent l’expressivité des émotions (Blanchard & Cohen, 2006), l’augmentation des traits affectifs négatifs (Horan et al., 2008), les troubles de la perception des signaux socioaffectifs (Hoekert, Kahn, Pijnenborg & Aleman, 2007) et une plus forte expérience des émotions négatives (Cohen & Minor, 2010). Depuis quelques années, des études en neuro-imagerie fonctionnelle ayant analysé les substrats neuronaux associés à la perception émotionnelle chez les patients atteints de schizophrénie, ont souligné des altérations marquées, principalement observées en ce qui concerne l’amygdale, du CPFm et de l’aire postérieure (Taylor et al., 2012). De plus, chez les patients schizophrènes, il est fréquent de retrouver une activation des régions limbiques et préfrontales réduites pour les stimuli déplaisants, comparativement aux stimuli neutres, lors d’une tâche de mémoire émotionnelle (Lakis et al., 2011 ; Schneider et al., 1998 ; Takahashi et al., 2004). De prime abord, ceci pourrait indiquer un seuil de sensibilité différent à l’activation et à la valence émotionnelle chez les patients. Par contre, on a très peu tenté de séparer les contributions de la valence émotionnelle et de la charge activatrice de ces stimuli, surtout dans un protocole utilisant la technique des potentiels évoqués cognitifs.
Les composantes de potentiels évoqués cognitifs utilisées en schizophrénie
Les potentiels évoqués cognitifs (PÉC) sont des fluctuations de voltage électro-cortical dans un segment du tracé EEG et qui se manifestent en réponse à un événement cognitif. Pour améliorer le rapport signal-bruit, nous bénéficions de procédures de moyenne et de filtrage permettant de faire émerger le signal de l’activité neuronale associée à des composantes permettant d’y associer des processus cognitifs, moteurs ou sensoriels[1] (Luck, 2005). Pour les besoins du présent travail, il sera question des composantes N200, P300 et LPC qui sont généralement présentes dans une tâche de mémoire épisodique visuelle.
L’évaluation précoce des émotions et la composante N200 dans la schizophrénie
La composante N200 atteint son amplitude maximale à environ 200 millisecondes (variant entre 150 et 400 ms environ) suivant l’apparition d’un stimulus. À ce titre, une étude a montré des effets intéressants durant la présentation d’images contrastant la valence et l’activation émotionnelle. Dans ce contexte, les travaux de Champagne (2014) ont étudié les potentiels évoqués chez les patients atteints de schizophrénie. Les résultats indiquent que, dans le groupe témoin, l’amplitude de la N200 était plus latéralisée sur l’hémisphère droit, alors qu’il n’y avait pas cette latéralisation chez les patients atteints de schizophrénie. Par ailleurs, bien que les problèmes de mémoire non émotionnelle dans la schizophrénie soient assez bien documentés et que les problèmes associés à la perception des émotions soient aussi étayés, aucune étude n’a étudié les effets combinés de la mémoire en fonction des émotions, dans une population clinique bien sélectionnée. Par ailleurs, peu d’efforts furent investis pour approfondir la compréhension des substrats électro-corticaux de la mémoire épisodique émotionnelle chez les personnes atteintes de schizophrénies.
Dans un protocole de mémoire épisodique, la N200 refléterait les processus précoces associés à la familiarité (Pickering & Schweinberger, 2003) où l’amplitude de cette composante serait plus ample aux stimuli inconnus qu’aux stimuli connus en réponse à la présentation d’images photographiques (Lavoie & O’Connor, 2013). Toutefois, peu d’études ont été centrées sur la N200 dite « émotionnelle » dans la modalité visuelle et de surcroît dans un protocole de mémoire épisodique et encore moins avec un groupe atteint de schizophrénie. De façon plus générale, la composante N200, en réponse à des stimuli auditifs, est une composante connue pour présenter une déflexion d’amplitude réduite chez les individus atteints de schizophrénie (O’Donnell et al., 1994 ; Salisbury, O’Donnell, McCarley, Shenton & Benavage, 1994). Bien entendu, il est difficile d’inférer ces données d’une population clinique à un quelconque problème de traitement de la familiarité lié à la N200 dans la schizophrénie. Néanmoins, certaines données intéressantes permettent de soulever l’hypothèse d’une altération de cette composante. Ainsi, la N200 est aussi corrélée avec des anomalies neuro-anatomiques, comme une réduction du volume du lobe temporal médian et celui de la région néocorticale dans la schizophrénie (Egan et al., 1994 ; O’Donnell et al., 1993). De plus, la latence de la N200 a été corrélée avec les symptômes négatifs chez les personnes atteintes de schizophrénie (Stefansson & Jonsdottir, 1996).
La composante P300 et la réponse émotionnelle dans la schizophrénie
La composante P300 est une déflexion positive prenant forme entre 250 et 500 ms après la présentation du stimulus (Polich, 2007). L’une des premières études montrait que l’amplitude de la P300 était réduite dans le groupe atteint de schizophrénie, en comparaison à un groupe contrôle (Roth & Cannon, 1972), ce qui a été confirmé par d’autres études (Hill & Weisbrod, 1999). Les paradigmes utilisés joueraient un rôle important dans la compréhension de ces facteurs, pour l’utilisation de cet indice en recherche clinique (Jeon & Polich, 2003). De plus, ces anomalies ne seraient pas entièrement dues à une altération des processus contrôlés, mais seraient plutôt la conséquence de déficits cognitifs relativement précoces dans la chaîne de traitement de l’information (Alain, Bernstein, Cortese, Yu & Zipursky, 2002), suggérant que les patients arrivent à traiter cette information, mais que celle-ci serait dégradée en raison d’un mauvais décodage précoce (Foxe, Doniger & Javitt, 2001). Ceci souligne l’importance de tenir compte des composantes en amont, qui précèdent la P300. Pour ce qui est de la réponse aux émotions de la composante P300, les résultats montrent que, chez les sujets contrôle, les images à valences déplaisantes génèrent des composantes P300 de plus grande amplitude que les images plaisantes (An et al., 2003). À l’inverse, les patients schizophrènes obtenaient des amplitudes P300 plus faibles pour les images déplaisantes, en comparaison aux images plaisantes, suggérant que ces patients présentent d’importantes altérations dans la reconnaissance ou le traitement des émotions déplaisantes (An et al., 2003 ; Champagne, Mendrek, Germain, Hot & Lavoie, 2014). Un point intéressant réside dans le fait que cette réduction de la P300 antérieure était aussi corrélée positivement avec les symptômes négatifs de psychose.
La composante LPC et la réponse émotionnelle dans la schizophrénie
Dans un protocole de mémoire épisodique, cette composante tardive, aussi nommée Late Positive Component (LPC) suit normalement la composante P300, entre 500 et 1000 ms post-stimulus et refléterait les processus de récollection consciente (Curran & Cleary, 2003 ; Rugg et al., 1998 ; Wilding & Rugg, 1996). Sur le plan émotionnel, peu d’études ont documenté la sensibilité de cette composante aux dimensions de valence ou d’activation émotionnelle. Glaser et al. (2012) ont mis en évidence que la valence émotionnelle influencerait l’effet de mémoire dans des fenêtres relativement précoces (N200 et P300), alors que l’activation émotionnelle aurait un impact plus tardif dans la fenêtre de la LPC. D’autres études avaient précédemment trouvé des effets similaires de mémoire de latences précoces associées à la valence émotionnelle, alors que des effets plus tardifs étaient associés à l’activation émotionnelle (Cuthbert, Schupp, Bradley, Birbaumer & Lang, 2000 ; Diedrich O., 1997 ; Dolcos & Cabeza, 2002 ; Van Strien, Langeslag, Strekalova, Gootjes & Franken, 2009). Une hypothèse avancée par Van Strien et al. (2009) souligne que la valence aurait un impact sur l’effet de mémoire épisodique (effet connu inconnu), et qu’il serait un corollaire d’un processus de reconnaissance rapide provenant des aires préfrontales. À l’inverse, l’activation émotionnelle interagirait plutôt avec un processus d’encodage soutenu et plus tardif dans le temps. Pour ce qui est des effets de la LPC en fonction de la mémoire émotionnelle, chez les patients schizophrènes, une étude récente a montré une réduction marquée d’amplitude de la LPC en réponse aux images à forte activation émotionnelle suivant des descripteurs neutres (Horan, Wynn, Kring, Simons & Green, 2010 ; Strauss et al., 2013), suggérant une possible incapacité des patients à réguler une activation émotionnelle plus forte.
Objectifs et hypothèses
L’originalité de la présente étude réside dans le fait d’examiner la mémoire émotionnelle, alors que la vaste majorité des études de potentiels évoqués cognitifs, dans la schizophrénie, se concentrent soit sur les émotions, soit sur la mémoire épisodique, mais pas sur l’interaction entre les deux. Dans le présent article, nous proposons donc une méthode adaptée pour l’étude du traitement émotionnel, soit le système International Affective Picture System (IAPS) enchâssé dans un protocole de mémoire épisodique en potentiels évoqués cognitifs. Les caractéristiques de l’IAPS se basent principalement sur les dimensions de valence et d’activation des stimuli et furent largement utilisées dans le domaine de l’électrophysiologie cognitive (Olofsson, Nordin, Sequeira & Polich, 2008).
En premier lieu, nous voulons approfondir de quelles façons la valence et l’activation émotionnelle vont affecter la mémoire épisodique des patientes atteintes de schizophrénie. Nous avons fait le choix d’un groupe de femmes, afin d’homogénéiser nos groupes en fonction du sexe, puisque les images IAPS furent standardisées séparément pour les femmes et les hommes. Une autre raison de choisir un groupe de femmes est la disponibilité de leurs bilans hormonaux complets afin de contrôler les fluctuations possibles liées aux cycles menstruels. De plus, l’âge d’apparition de la schizophrénie est habituellement plus tardif chez les femmes que chez les hommes (van der Werf et al., 2012), alors que l’âge (Addis, Leclerc, Muscatell & Kensinger, 2010) et le sexe (Tsai, McDowd, Tang & Su, 2012) influencent aussi les performances mnésiques.
Par ailleurs, les processus associés à la mémoire émotionnelle possèdent une chronométrie d’apparition relativement précoce et il est primordial d’utiliser les potentiels évoqués cognitifs qui permettront de suivre les étapes de traitement se déroulant à l’échelle de quelques centaines de millisecondes afin de cibler les processus mnésiques et émotionnels sur une ligne temporelle fine.
Sur le plan topographique, nous proposons l’hypothèse que l’activité des régions frontales associées à certains types de stimuli émotifs sera généralement affectée dans la population clinique, et ce, particulièrement en réponse aux stimuli à fortes activations émotionnelles. Sur le plan du décours temporel, et en fonction des composantes à l’étude, il demeure hasardeux d’émettre une hypothèse explicite sur la composante N200, puisque relativement peu d’études de potentiels évoqués ont fait état d’une atteinte fonctionnelle touchant cette composante dans un protocole de mémoire émotionnelle avec une population de patients schizophrènes. Toutefois, si nous nous basons sur les modèles de mémoire épisodique associant le concept de familiarité à une activité frontale précoce, nous pouvons raisonnablement proposer l’hypothèse que l’amplitude de la N200 frontale sera diminuée dans toutes les conditions émotionnelles. Par ailleurs, les patients schizophrènes devraient avoir un effet de mémoire réduit pour la P300 frontale dans les conditions à valence émotionnelle déplaisante, en comparaison aux images plaisantes, alors que chez les contrôles, les images à valence émotionnelles déplaisantes vont plutôt générer pour la composante P300 des effets de mémoire de plus grandes amplitudes que les images plaisantes. Plus tardivement, les données disponibles nous conduisent à prédire que les patientes schizophrènes auront un effet de mémoire LPC réduit en réponse aux images à forte activation, et ce, toujours dans les régions frontales.
Méthode
Participants
Seize patientes schizophrènes, répondant aux critères diagnostics du DSM-IV-TR pour la schizophrénie (APA, 2000) dans une phase stable de leur maladie, furent appariés à 17 participantes contrôles sur la base de l’âge, de la dominance manuelle (droitière ou gauchère) selon le test d’Edinburgh (Oldfield, 1971) et le statut socio-économique des parents (Classification nationale des professions, 2001). Toutes les patientes ont été réévaluées par des psychiatres expérimentés, et furent affectées au groupe de recherche (tableau 1). Les troubles de l’humeur, les troubles schizos affectifs et schizophréniformes ont été exclus. Les participantes du groupe témoin ont passé une entrevue clinique de dépistage avec l’édition pour les non patients du Structured Clinical Interview for DSM Disorders (SCID) (First, Spitzer, Gibbon & Williams, 1996). La gravité des symptômes a été évaluée selon l’échelle de syndrome positif et négatif (PANSS) (Kay, Fiszbein & Opler, 1987). Le début de la maladie a été défini comme étant la date de première consultation psychiatrique. Toutes les patientes avaient reçu au moins un antipsychotique atypique (10 des patientes ont reçu un seul type d’antipsychotique et 6 en ont reçu deux ; clozapine : n = 9, dose moyenne = 377,78 mg ± 119,53 mg ; olanzapine : n = 4, dose moyenne = 15 mg ± 7,07 mg ; rispéridone : n = 7, dose moyenne = 4 mg ± 2,08 mg ; quétiapine : n = 2, dose moyenne = 318,75 mg ± 326,2 mg). Toutes les doses d’antipsychotiques ont été calculées en équivalence chlorpromazine (Woods, 2003).
Les critères d’exclusion générale de l’étude comprenaient l’âge en dessous de 18 ou plus de 45 ans, un passé ou un présent de trouble neurologique ou psychiatrique concomitant à l’axe-1, l’alcoolisme ou la toxicomanie, le non-respect des procédures de test et une vision anormale non corrigée. Ceux qui n’ont pas pu recevoir de prise de sang la même journée que l’enregistrement EEG étaient systématiquement exclus (n = 14 exclusions). La capacité des patientes atteintes de schizophrénie à donner un consentement éclairé a été établie en utilisant les lignes directrices de l’Association canadienne de psychiatrie (Arboleda-Florez, 1997) et l’étude a été approuvée par les comités d’éthique de la recherche locaux (IUSMM et Regroupement Neuroimagerie Québec).
Tableau 1
Comparaison des moyennes et écarts-types (parenthèses) des caractéristiques démographiques et cliniques

* ; p < ,05 ** p < ,005
WAIS : Weschler Adult Intelligence Scale ; PANSS : Positive and Negative Syndrome Scale, SSE= Statut socioéconomique ; NS : non significatif
Cadre expérimental
On demandait à toutes les patientes de se présenter au laboratoire pour l’étude lors des jours 4 à 8 ou 16 à 26 de leurs cycles menstruels. Ceci avait pour objectif de vérifier les différences hormonales des femmes tout au long de leur cycle au moment où elles se présentaient au laboratoire, puisque ces facteurs influencent l’EEG (Champagne et al., 2014) et l’activité métabolique (Champagne et al., 2012). Mis à part la prolactine, aucune différence significative entre les niveaux d’hormones (estrogène, progestérone et testostérone) entre les femmes du groupe contrôle et les femmes du groupe schizophrène n’a été rapportée (voir tableau 2). Les participantes étaient assises dans une chaise réglable, placée devant l’écran d’ordinateur à une distance de 90 cm, calculée à partir du nez vers le centre de l’écran (angle de 5 degrés). Les images en format JPEG (1280 × 960 pixels ; maximum 16,7 millions de couleurs) ont été présentées en séquence sur un écran plat 19’’ (NEC AccuSync LCD 92vx), avec un taux de rafraîchissement de 60 Hz, pour une durée fixe de 3000 ms et les intervalles entre les essais ont été fixés à 1000 ms, au cours de laquelle une image de damier rouge et blanc apparaissait (# IAPS 7182).
Tableau 2
Comparaison des moyennes et écarts-types (parenthèses) du bilan hormonal
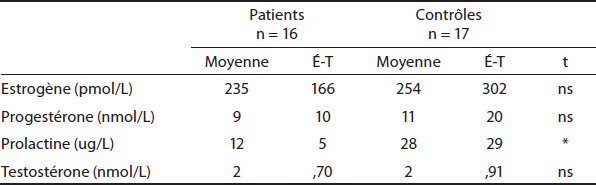
* p < ,05 ; ** p < ,005
Sélection des stimuli et protocole de mémoire épisodique[2]
Le protocole de présentation des images était aussi utilisé dans le cadre d’un protocole de mémoire épisodique ayant eu lieu avec des participants contrôles (Glaser etal., 2012). Ce protocole mnésique comportait une section étude suivie d’une section rappel épisodique. Le matériel émotionnel utilisé était des photographies d’images sélectionnées à partir du système international d’images affectives, l’International Affective Picture System – IAPS (Lang, Bradley & Cuthbert, 2008), une collection standardisée d’images recueillies à partir d’une variété de catégories émotionnelles et sémantiques. Un total de 200 images a été sélectionné et réparti en quatre catégories équivalentes en fonction de leur valence et de leur niveau d’intensité. La sélection des images a été réalisée sur la base de l’évaluation normative IAPS standardisée pour les femmes. Les quatre groupes d’images comptaient 50 photos chacun (1 – valence plaisante ou positive et avec une forte intensité (ex : photos érotiques), 2 – valence plaisante avec faible intensité activatrice (ex : chatons), 3 – valence déplaisante ou négative avec une forte intensité activatrice (ex : des images de guerre), 4 – valence déplaisante avec une faible intensité activatrice (ex : un enfant qui pleure).
La moitié des images est présentée une seule fois en retest (condition inconnue : 100 images) et l’autre moitié présentée une deuxième fois (condition connue : 100 images), sont divisées en 4 catégories de 50 images par catégorie présentées dans un ordre semi-aléatoire. Dans chaque catégorie émotionnelle, les images contenaient certains attributs de base similaires (des humains, des animaux, des objets inanimés ou des paysages) afin de préserver la cohérence entre les conditions connues et inconnues.
Enregistrement de l’EEG et extraction des potentiels évoqués cognitifs (ERP)
L’EEG a été enregistré à partir de 60 électrodes en Ag/AgCl fixées dans un casque en lycra (Electrode Arrays, Sands Research, Inc), placées selon l’orientation standard pour la position des électrodes par l’American EEG Society (Pivik et al., 1993). Seulement 24 électrodes furent conservées, afin d’étudier les régions d’intérêts selon les localisations frontales (F1, F2, F3, F4, F5 et F6 FC1, FC2, FC3, FC4, FT7 et FT8) et pariétales (C1, C2, C3, C4, C5 et C6, P1, P2, P3, P4, P5 et P6). Toutes les électrodes ont été référencées aux mastoïdes et leurs impédances ont été maintenues en dessous de 5 kΩ. Les électro-oculogrammes (EOG) ont été enregistrés en utilisant quatre électrodes bipolaires externes en étain de 9 mm (EOG horizontal sur chacune des tempes et pour l’EOG vertical aux points infra et supra-orbitaires de l’oeil gauche). Le signal était acquis par un amplificateur bioélectrique digital DBPA-1 (Sensorium, Charlotte, VA) avec un gain calibré (± 5000) et une bande passante comprise entre 0,01 et 30 Hz, ainsi qu’un taux d’échantillonnage de 500 Hz. Les corrections oculaires étaient ensuite réalisées sur le signal continu afin de retirer tout artéfact oculaire pouvant contaminer l’EEG. Les signaux étaient ensuite moyennés par rapport au stimulus (100 ms avant et jusqu’à 1000 ms après le stimulus avec bornes d’exclusion à + 100 uV et - 100 uV). La quantité minimale d’essais tolérée dans une condition donnée était de 16 essais libres d’erreurs et d’artéfacts afin d’être considérés valides, ce qui correspond à des études antérieures, avec un protocole de mémoire épisodique d’images photographiques émotionnelles (Glaser et al., 2012). Les composantes furent analysées à partir des pics d’amplitudes maximales dans trois fenêtres de temps sensibles aux aspects émotionnels du traitement des stimuli visuels (Kissler, Herbert, Winkler & Junghofer, 2009 ; Schupp, Junghofer, Weike & Hamm, 2003 ; Stewart et al., 2010), dont la N200 (200-400 ms), la P300 (300-500 ms) et la LPC (500-1000 ms). Ces trois composantes furent analysées en groupant les régions antérieures vs postérieures.
Analyse de données et statistiques
Les variables dépendantes d’intérêt, soit l’amplitude maximale, baseline-to-peak, pour chacune des composantes des PÉC, ont été analysées séparément à l’aide d’analyses de variance à mesures répétées ANOVA (SPSS® – Win 20). Les données comportementales (évaluation subjectives des images et les temps de réaction durant la tâche) et électrophysiologiques (PÉC) ont été soumises à une MANOVA à mesures répétées (SPSS® – Win 20) avec comme facteur intrasujet, la condition mémoire (deux niveaux : connus/inconnus), la valence (deux niveaux : plaisant/déplaisant) et l’activation (deux niveaux ; faible/élevée). Les facteurs inter-groupes étaient composés du groupe (deux niveaux : schizophrènes/contrôles). Des facteurs additionnels furent analysés en incluant les niveaux intrasujets hémisphériques (2 niveaux : gauche/droite). Pour notre analyse, les électrodes situées en frontal, en frontal-central et en central ont été regroupées pour représenter les régions antérieures, alors que les électrodes situées sur la région pariétale furent analysées séparément pour représenter la région postérieure. Les analyses ont été effectuées séparément sur chacune des trois fenêtres temporelles (N200, P300 et LPC) et chacune des régions cérébrales (frontales et pariétales). Pour toutes les analyses, le niveau de signification a été fixé à 5 % (bidirectionnel) et une correction epsilon (Huynd-Feldt) était appliquée lorsque nécessaire.
Résultats
Évaluations subjectives des images émotionnelles
Les résultats des évaluations subjectives révèlent des moyennes de moins d’un écart-type par rapport à la norme de l’IAPS des femmes contrôles (tableau 3). Les analyses des scores de l’évaluation subjective de la valence montrent un effet principal de valence (F [1,31] = 231,99 ; p < ,001) ainsi qu’une interaction valence par activation (F [1,31] = 29,72 ; p < ,001), ce qui signifie que les images déplaisantes à activations élevées sont évaluées plus déplaisantes, alors que l’activation a peu d’impact sur l’évaluation de la valence plaisante. Il n’y a pas d’effet de répétition (effet connu inconnu) ou de différences de groupe significatives dans l’évaluation, ce qui montre que les stimuli connus ou inconnus ne diffèrent pas au niveau de l’évaluation de la valence émotionnelle et que les patientes évaluent cette dimension de façon équivalente aux contrôles. En deuxième lieu, les scores d’évaluations de l’activation émotionnelle montrent un effet principal d’activation (F [1,31] = 99,40 ; p < ,001), ce qui dénote que les évaluations de l’activation sont conformes à ce qui était attendu. Il n’y a pas d’effet de répétition (effet connu inconnu) ou de groupe, ce qui démontre que les stimuli connus ou inconnus ne diffèrent pas au niveau de l’évaluation de l’activation et que dans l’ensemble, il n’y a aucune différence de groupe significative dans l’évaluation de la valence ou de l’activation émotionnelle (tableau 3).
Tableau 3
Comparaison des évaluations subjectives de la valence et de l’activation émotionnelle en fonction des types de réponses en mémoire (connus vs inconnus) dans les deux groupes, ainsi que pour la norme IAPS pour les femmes

Valence : Évaluation de la valence émotionnelles selon une échelle de 1 à 9 (1 = très déplaisant | 5 = neutre | 9 = très plaisants)
Activation : Évaluation de l’activation émotionnelle selon une échelle de 1 à 9 (1= activation très faible à 9 = activation très forte)
Temps de réaction
Les temps de réaction montrent des effets principaux de valence (F [1,31] = 4,60 ; p < ,05) et d’activation (F [1,31] = 34,66 ; p < ,001), ainsi qu’un effet d’interaction valence x activation (F [1,31] = 4,63 ; p < ,05). Ainsi, les images à activations élevées ralentissent significativement les temps de réaction par rapport aux images à activations faibles. Toutefois, ce délai associé aux images à activations élevées est plus important dans la condition à valence émotionnelle plaisante que dans la condition déplaisante. Nous notons aussi un effet d’interaction de mémoire x activation (F [1,31] = 40,80 ; p < ,001) ce qui dénote un effet d’activation émotionnelle plus important lorsque les stimuli sont connus (89 ms) que lorsqu’ils sont inconnus (4 ms). Outre un ralentissement non significatif aux images déplaisantes, la valence n’a toutefois pas d’impact sur la condition de mémoire et nous ne notons aucune différence de groupe significative (tableau 4).
Tableau 4
Comparaison des temps de réaction en millisecondes (erreur-type) en réponse à la valence (plaisantes et déplaisantes) et à l’activation (élevées et faibles) émotionnelle en fonction des types de réponses en mémoire (connus vs inconnus) dans les deux groupes

Note : Pl = valence plaisantes ; Dpl = valence déplaisantes ; É = activations élevées ; F = activations faibles
Électrophysiologie
N200 frontale : L’amplitude de la N200 frontale montre un effet principal de mémoire (F F [1,32] = 9,70 ; p < ,005), ce qui dénote une amplitude trois fois plus importante dans la condition inconnue (- 1,6 uV) que dans la condition connue (- 0,5 uV). Pour ce qui est de l’effet des émotions, il y a un effet de valence (F [1,32] = 7,23 ; p < ,05), ainsi qu’un effet d’interaction valence x activation (F [1,32] = 36,43 ; p < ,001), ce qui met en évidence une amplitude de la N200 plus importante en réponse à la présentation d’images déplaisantes et à fortes activations (- 2,74 uV), par rapport à la condition plaisante à forte activation (0,28 uV). Une interaction quadruple de groupe x mémoire x valence x hémisphères (F [1,32] = 8,36 ; p < ,01) est aussi relevée. Ainsi, cette interaction complexe dénote une augmentation de l’effet de mémoire dans l’hémisphère droit en réponse aux stimuli déplaisants, chez les patientes schizophrènes. Pour ce qui est du groupe contrôle, il y a aussi une augmentation de cet effet de mémoire dans l’hémisphère droit, mais cette fois, en réponse aux stimuli plaisants (figure 1).
Figure 1

Amplitude de la N200 frontale en réponse aux conditions de mémoire épisodique connu (cercle noir) et inconnu (triangle blanc) en fonction des groupes et des conditions de valence émotionnelle. En dépit d’une N200 frontale généralement réduite, les patientes démontrent une augmentation de l’effet de mémoire N200 dans l’hémisphère droit par rapport à l’hémisphère gauche, en réponse aux stimuli déplaisants. Pour ce qui est du groupe contrôle, il y a aussi une augmentation de cet effet de mémoire dans l’hémisphère droit, mais cette fois, en réponse aux stimuli plaisants.
N200 pariétale : L’amplitude de la N200 pariétale montre encore un effet principal de mémoire (F [1,32] = 11,02 ; p < ,005), ce qui dénote une amplitude plus importante dans la condition inconnue (5,40 uV) que dans la condition connue (4,74 uV). Les aspects associés à la valence ou à l’activation émotionnelle n’ont aucun effet significatif sur la N200 pariétale. Il y a aussi un effet principal de groupe (F [1,32] = 5,13 ; p < ,05), ce qui dénote que les patientes schizophrènes ont généralement une amplitude N200 moins importante que celle des contrôles. Toutefois une interaction groupe x mémoire (F [1,32] = 7 18 ; p < ,05) souligne que l’effet de mémoire sur la N200 pariétale est généralement plus important chez les contrôles (1,20 uV) que chez les patientes (0,12 uV), peu importe la condition de valence ou d’activation émotionnelle.
P300 frontale : L’amplitude de la P300 frontale montre un effet principal de mémoire (F F [1,32] = 12,36 ; p < ,005), ce qui dénote une amplitude plus importante dans la condition connue (- 1,23 uV) que dans la condition inconnue (0,28 uV). Pour ce qui est de l’effet des émotions, il y a un effet de valence (F [1,32] = 9,25 ; p < ,01), un effet d’activation (F [1,32] = 4,70 ; p < ,05), ainsi qu’un effet d’interaction valence x activation (F [1,32] = 23,68 ; p < ,001) et une interaction mémoire x valence x activation (F [1,32] = 7,79 ; p < ,01) ce qui met en évidence que l’effet de mémoire épisodique est plus important en réponse à la présentation d’image à la fois déplaisante et à faible activation (3,09 uV), par rapport à la condition déplaisante à forte activation (0,02 uV), alors que cet effet de mémoire n’est pas significativement affecté par l’activation, dans la condition plaisante. Par ailleurs, nous notons un effet principal de groupe ([1,32] = 5,15 ; p < ,05) qui dénote une composante P300 frontale globalement plus élevée dans le groupe patient (1,75 uV) que dans le groupe contrôle (- 2,70 uV). Une interaction mémoire x hémisphères (F [1,32] = 4,78 ; p < ,05) et une interaction quadruple de groupe x mémoire x valence x hémisphères (F [1,32] = 9,01 ; p < ,05) est aussi relevée. Ainsi, cette interaction complexe dénote une augmentation de l’effet de mémoire dans l’hémisphère droit (2,72 uV) en réponse aux stimuli déplaisants, chez les patientes schizophrènes. Pour ce qui est du groupe contrôle, il y a aussi une augmentation de cet effet de mémoire dans l’hémisphère droit (3,67 uV), mais cette fois, en réponse aux stimuli plaisants (Figure 2).
Figure 2

Amplitude de la P300 frontale en réponse aux conditions de mémoire épisodique connu (cercle noir) et inconnu (triangle blanc) en fonction des groupes et des conditions de valence émotionnelle. Les patientes démontrent une augmentation de l’effet de mémoire P300 dans l’hémisphère droit, uniquement en réponse aux stimuli déplaisants. Le groupe contrôle démontre un plus grand effet de mémoire dans l’hémisphère droit en réponse aux stimuli plaisants.
P300 pariétale : L’amplitude de la P300 pariétale montre un effet principal de mémoire (F [1,32] = 10,36 ; p < ,005), et une interaction mémoire x groupe (F [1,32] = 6,54 ; p < ,05) ce qui dénote que l’effet de mémoire épisodique est plus important pour le groupe contrôle (2 uV) que pour le groupe de patients (0,2 uV). Pour ce qui est de l’effet des émotions, il y a un effet de valence (F [1,32] = 8,29 ; p < ,05), un effet d’activation (F [1,32] = 8,80 ; p < ,01), ainsi qu’une interaction valence x activation (F [1,32] = 10,08 ; p < ,001) ce qui met en lumière une amplitude de la P300 plus importante en réponse à la présentation d’image à la fois plaisante et à forte activation (7,45 uV), par rapport à la condition déplaisante à forte activation (5,80 uV). Il n’y avait pas de différence d’amplitude en réponse aux stimuli avec une faible charge activatrice et les deux groupes exprimaient une P300 pariétale similaire en réponse à ces conditions expérimentales (figure 4).
Figure 3

Illustration des potentiels évoqués cognitifs frontaux de l’hémisphère gauche et droit moyennés pour les images plaisantes (panneau A) et déplaisantes (panneau B). Les patientes schizophrènes dénotent une N200 réduite, alors que la P300 et la LPC sont généralement plus amples que chez les contrôles.
LPC frontale : L’amplitude de la LPC frontale montre un effet d’interaction mémoire x groupe (F [1,32] = 7,25 ; p < ,05), ce qui dénote un effet de mémoire plus important pour le groupe contrôle (2,19 uV) que pour le groupe de patientes (0,56 uV). Toutefois, nous retrouvons aussi une interaction mémoire x activation (F [1,32] = 11,15 ; p < ,05), mémoire x valence x activation (F [1,32] = 5,69 ; p < ,05) et une interaction à quatre niveaux, groupe x mémoire x valence x activation (F [1,32] = 4,70 ; p < ,05). Ainsi, chez le groupe contrôle, l’effet de mémoire épisodique (c.-à-d. plus grande amplitude pour la condition connue qu’à la condition inconnue) est plus important en réponse aux stimuli plaisants à forte activation qu’aux stimuli plaisants à faible activation, alors que cet effet de mémoire disparaît pour les stimuli déplaisants à forte activation. Dans le groupe de patientes, nous retrouvons un effet de mémoire inverse, c’est-à-dire un effet d’amplitude plus important pour les stimuli inconnus (1,91 uV) que pour les stimuli connus (- 2,18 uV), et ce, pour les stimuli à forte activation émotionnelle, tant pour les stimuli à valence plaisante que déplaisante (figure 3).
Figure 4

Illustration des potentiels évoqués cognitifs pariétaux de l’hémisphère gauche et droit moyennés pour les images plaisantes (panneau A) et déplaisantes (panneau B). L’effet de mémoire sur la N200 pariétale est généralement réduit chez les patientes schizophrènes en comparaison au groupe contrôle. Plus loin dans le décours temporel, le groupe de patientes dénote une amplitude de la composante P300 significativement plus importante, mais l’effet de mémoire épisodique est toutefois réduit dans ce groupe, et ce, dans toutes les conditions émotionnelles.
LPC pariétale : L’amplitude de la LPC pariétale montre un effet d’interaction groupe x mémoire (F [1,32] = 5,67 ; p < ,05), ce qui dénote un effet de mémoire plus important pour le groupe contrôle (1,11 uV) que pour le groupe de patientes (0,65 uV). Pour ce qui est de l’effet des émotions, il y a une interaction mémoire x valence x activation (F [1,32] = 4,73 ; p < ,05) qui dénote que l’effet de mémoire épisodique est plus important dans la condition déplaisante à faible activation que dans la condition déplaisante à forte activation émotionnelle, alors qu’il n’y a pas d’effet de mémoire dans la condition plaisante. Ce patron d’effet est similaire dans les deux groupes (figure 4).
Discussion
Étant donné l’état des connaissances concernant l’activité cérébrale, l’état affectif et le traitement des émotions des personnes atteintes de schizophrénie, nous avions suggéré que l’activité des régions frontales serait généralement moindre en réponse aux stimuli émotionnels. Nous avions formulé l’hypothèse d’une diminution de la N200 au niveau frontal chez les patientes. Par ailleurs, nous avions proposé que les patientes schizophrènes aient un effet de mémoire réduit pour la P300 frontale dans les conditions à valence émotionnelle déplaisante, en comparaison aux images plaisantes. Plus tardivement, nos hypothèses prédisaient que les patientes schizophrènes auraient un effet de mémoire LPC réduit en réponse aux images à forte activation, et ce, dans les régions frontales.
Notre paradigme a permis de dissocier les effets liés à la valence et à l’activation émotionnelle et de quantifier leurs impacts sur la mémoire. En premier lieu, les temps de réaction permettent de voir qu’il y a un délai significatif dans les réponses, lorsque les images sont à activation élevée, et ce, particulièrement dans la condition à valence émotionnelle plaisante, confirmant ainsi les résultats de recherches passées (Glaser et al., 2012 ; Maratos, Allan & Rugg, 2000 ; Windmann & Kutas, 2001). Par contre, les temps de réaction n’ont pas fait ressortir les effets de groupes, ce qui implique qu’au niveau des réponses comportementales, les patientes réagissent comme les contrôles. De même, nous notons aussi des performances équivalentes pour l’évaluation subjective de la valence et de l’activation émotionnelles des photographies.
Impact des émotions et de la mémoire épisodique en région frontale
Au niveau de la N200 frontale, nous notons une sensibilité plus importante pour la valence, que pour l’activation émotionnelle. De plus, certaines différences de groupe semblent ressortir en fonction de la mémoire et de la valence. Premièrement, la N200 frontale est significativement sous activée pour les patientes schizophrènes. Ensuite, des effets hémisphériques importants permettent de bien discriminer les groupes. Ainsi, tant pour la N200 que pour la P300 frontale, la magnitude de l’effet de mémoire épisodique observée chez les patientes schizophrènes est augmentée spécifiquement dans l’hémisphère frontal droit en réponse aux stimuli déplaisants, alors que l’augmentation de cette asymétrie en faveur de l’hémisphère droit est présente pour les stimuli plaisants chez les participantes contrôle. De plus, nos résultats montrent que les stimuli à valences émotionnelles plaisantes élicitent un effet de mémoire épisodique plus important dans l’hémisphère gauche chez les patientes schizophrènes, ce qui recoupe bien les résultats de Champagne et al. (2014).
Dans le contexte d’une tâche de mémoire épisodique, un effet N200 relativement précoce (300 à 500 ms) observé en région frontale bilatérale se produit lorsque l’accès à l’information perceptuelle et conceptuelle lié à l’item à reconnaître (item test) est facilité (Curran, 1999 ; Rugg et al., 1998a ; Ullsperger, Mecklinger, Matthes-von Cramon & Yves, 2000). Cette composante frontale précoce est sensible aux changements contextuels dans la reconnaissance des visages aussi liée à la familiarité (Guillaume & Tiberghien, 2001 ; Hintzman, Caulton & Levitin, 1998). Une amplitude de la N200 frontale de l’hémisphère droit a été précédemment identifiée, dans un paradigme de mémoire épisodique (Curran, 2000) et plusieurs recherches ont montré qu’elle refléterait l’évaluation de la familiarité, plutôt que la récollection consciente (Curran, 1999 ; Friedman et al., 2000 ; Rugg et al., 1998). Le concept de familiarité est défini comme la capacité d’évaluer la similitude conceptuelle entre un élément test (c.-à-d. connu) et toutes les informations de la liste d’étude en mémoire (Hintzman, Caulton & Levitin, 1998), alors que la récollection consciente implique la récupération des informations spécifiques aux éléments étudiés, tels que les attributs physiques, les informations associatives et contextuelles ou d’autres informations sources.
La N200 est relativement bien documentée dans les protocoles de mémoire, mais peu d’analyses de la N200 frontale ont permis de comprendre ses fonctions dans le contexte de rappel d’images émotionnelles. Toutefois, l’interprétation la plus fréquemment utilisée pour décrire les effets sur la N200 frontale est que l’attention serait orientée par les qualités perceptuelles contenues dans la valence des stimuli, pour un processus émotionnel ultérieur de haut niveau (Dolcos & Cabeza, 2002 ; Schupp, Junghofer, Weike & Hamm, 2004). Il est intéressant de constater que selon une recension exhaustive, la valence des stimuli est reconnue pour influencer la latence et l’amplitude de la composante N200 (Olofsson et al., 2008). Selon ces études, cette composante a été localisée dans le cortex cingulaire antérieur au moyen d’algorithmes d’analyses de sources génératrices (Carretie, Hinojosa, Martin-Loeches, Mercado & Tapia, 2004 ; Carretie, Mercado, Hinojosa, Martin-Loeches & Sotillo, 2004). Transposée aux résultats de nos patientes, la N200 pourrait suggérer une altération de la familiarité, spécifiquement aux stimuli déplaisants, dans le contexte d’une tâche de mémoire épisodique, tel que reflété par la N200 frontale. Ceci pourrait indiquer une altération du cortex cingulaire antérieur, récemment documentée avec cette population, et avec une tâche émotionnelle semblable (Benedetti et al., 2011).
Plus tard dans le décours temporel, la P300 frontale montre une sensibilité à la fois pour la valence et pour l’activation émotionnelle. Nos résultats ont permis de souligner que pour cette composante P300 frontale, il y a une augmentation significative de l’effet de mémoire dans l’hémisphère droit en réponse aux stimuli déplaisants, chez les patientes schizophrènes. Fait intéressant, l’effet de mémoire observé à l’hémisphère gauche chez les contrôles pour les stimuli plaisants est absent chez les patientes schizophrènes. L’amplitude de la LPC frontale montre, quant à elle, des effets de mémoire impliquant la valence et l’activation, mettant en lumière des différences de groupes plus importantes lors de présentations d’image à valence plaisante à forte activation. De façon plus précise, l’effet de mémoire épisodique serait plus important en réponse aux stimuli plaisants à forte activation, où le groupe de patientes aurait un effet de mémoire inversé par rapport aux contrôles.
Au niveau fonctionnel, la P300 frontale serait générée lors de l’engagement des mécanismes de prétraitement partiellement attribués au lobe frontal (Comerchero & Polich, 1998, 1999 ; Polich, 2007). Comment ces mécanismes seraient-ils affectés par la valence des stimuli ? Le cortex préfrontal recevrait rapidement les informations du cortex visuel, qui rendrait possibles l’organisation et le déclenchement d’une réponse rapide (Bar, 2003 ; Bar et al., 2006). De plus, le gyrus orbitofrontal, le gyrus frontal supérieur, le gyrus cingulaire antérieur et l’amygdale s’avèrent être impliqués dans la régulation des émotions déplaisantes (Mak, Hu, Zhang, Xiao & Lee, 2009). Ces considérations peuvent suggérer que la régulation des émotions soit accomplie par la mise à jour dynamique de la valeur affective accordée à un stimulus (Ochsner, Bunge, Gross & Gabrieli, 2002), ce qui expliquerait l’implication de la P300 frontale lors de la visualisation des stimuli à valence déplaisante (Mak et al., 2009).
Si on considère l’activité plus tardive de la composante LPC frontale, plusieurs recherches ont proposé que les tâches de catégorisations plus difficiles engendrent des changements qualitatifs indexés par une composante LPC frontale reflétant certains processus survenant juste avant la catégorisation d’un stimulus (c.-à-d. avant le temps de réaction). À ce titre, Folstein & van Petten (2011) ont observé des effets d’amplitudes plus importants sur la LPC frontale, pour des stimuli plus difficiles à catégoriser, alors que d’autres recherches ont observé cette positivité frontale dans le contexte de tâches d’arithmétiques plus difficiles (Iguchi & Hashimoto, 2000 ; Ruchkin, Johnson, Canoune & Ritter, 1991 ; Ruchkin, Johnson, Mahaffey & Sutton, 1988). Puisque diverses composantes LPC frontales sont apparues dans différentes expériences avec différents contextes ou modalités, il demeure hardi de vouloir transposer ces résultats directement aux nôtres. Toutefois, nous pouvons avancer que les patientes schizophrènes vont procéder à un rappel épisodique des émotions par la mise à jour dynamique de la valence affective accordée à un stimulus, en activant différemment les circuits frontaux associés à la N200, à la P300 et pour la LPC. Les images émotionnelles seraient-elles plus difficiles à catégoriser pour les femmes atteintes de schizophrénie ? Il est difficile d’arriver à cette conclusion avec les données actuelles, car il faut rappeler, ici, qu’en dépit des multiples différences frontales observées, les patientes ont obtenu des performances similaires aux contrôles, puisqu’elles ont été en mesure d’évaluer les stimuli avec autant de précisions et avec des temps de réaction similaires aux contrôles. Dans un autre ordre d’idée, ceci pourrait mettre en lumière des mécanismes de compensations similaires à ceux documentés dans d’autres études avec des tâches de mémoire utilisées en imagerie cérébrale fonctionnelle, où des temps de réaction et des performances équivalentes ont été remarqués chez les patients avec schizophrénie, alors que des différences cérébrales importantes furent enregistrées (Kim etal., 2010 ; Whyte et al., 2006).
Les composantes pariétales en mémoire épisodique et les réponses émotionnelles chez les patientes schizophrènes
En résumé, les différences de groupes en régions pariétales apparaissent surtout pour la composante la plus précoce, soit la N200 pariétale. Dans cette fenêtre de temps, les patientes schizophrènes ont généralement un effet de mémoire réduit, en comparaison avec le groupe contrôle. Plus tard dans le traitement, l’amplitude de la P300 pariétale montre un impact de la mémoire, de la valence et de l’activation émotionnelle, sans effet de groupe significatif. Ainsi, nos résultats mettent en lumière que le groupe clinique exprime un effet de mémoire P300 pariétale réduit par rapport au groupe contrôle, mais il n’y a aucune interaction avec les conditions émotionnelles. Traditionnellement, la composante P300 pariétale serait associée au processus de prise de décision, à la probabilité d’apparition du stimulus (Duncan-Johnson & Donchin, 1982) et à la mise en contexte en mémoire de travail (Verleger, 2008). Cette composante pariétale apparaît cependant dans une variété de contextes et reflète probablement plusieurs classes de processus cognitifs, encore mal compris, qui se chevauchent temporellement et spatialement (Polich, 2007).
Par ailleurs, l’amplitude de la LPC pariétale montre un effet de mémoire plus important pour le groupe contrôle que pour le groupe de patientes, mais il n’y a pas d’interaction entre l’effet de mémoire, la valence et l’activation émotionnelle. Ceci implique que l’effet tardif (LPC) de mémoire en région pariétale est généralement réduit chez les femmes atteintes de schizophrénie, mais que la dimension émotionnelle n’a pas d’impact significatif à ce stade de traitement de l’information. Selon la distribution attendue et la latence, cette composante est analogue à la composante LPC classique observée au cours d’expériences portant sur la mémoire épisodique, avec des mots (Curran, 1999) et des images (Curran & Dien, 2003) non émotionnelles, associés aux processus de récollection consciente, indiquant aussi une sensibilité à la profondeur du traitement (Friedman, 1990 ; Rugg et al., 1998), qui suggérerait que l’information émotionnelle à valence déplaisante a un accès privilégié aux ressources cognitives (Dolcos & Cabeza, 2002) traitées par les zones corticales pariétales (Halgren et al., 1995 ; Paller, Gonsalves, Grabowecky, Bozic & Yamada, 2000 ; Rugg, Roberts, Potter, Pickles & Nagy, 1991). Ainsi, dans cette fenêtre temporelle, des voies neuronales semblables sont probablement actives durant la récollection consciente dans tous les types d’émotions, et cela, autant dans le groupe de patientes que chez les contrôles.
Limites de l’étude et orientations futures
Nos analyses comparaient les images connues par rapport aux images inconnues, mais nous n’avons pas pris en compte les erreurs ou les fausses alarmes. Dans la présente étude, nous avons tenu compte uniquement des bonnes réponses, tant pour le calcul de temps de réaction que pour les potentiels évoqués. Comme il s’agit d’une tâche de mémoire épisodique, une étude ultérieure pourrait approfondir en analysant séparément les réponses associées aux erreurs et aux bonnes réponses, à condition de générer suffisamment d’erreurs à la tâche, afin de permettre un signal comportant suffisamment de réponses moyennées. De plus, nous n’avons pas analysé les potentiels évoqués en réponse à la phase d’étude, lors de la première présentation des stimuli (phase étude). Dans une étude ultérieure, il serait important de tenir compte de cette phase étude (première exposition) pour voir si un phénomène d’amorçage émotionnel pourrait moduler les réponses observées à la phase test (deuxième exposition). Enfin, nous avons comparé deux groupes de femmes et il serait aussi judicieux d’ajouter une comparaison avec deux groupes d’hommes atteints ou non de schizophrénie.
Conclusion
En somme, les données montrent des différences entre les patientes et les contrôles sur plusieurs composantes temporellement et spatialement distinctes, associées aux effets émotionnels et, qui plus est, contribuent aux potentiels évoqués cognitifs observés. Ainsi, la majorité des effets liés à la schizophrénie sont retrouvés principalement au niveau des potentiels évoqués observés en régions frontales. En outre, ces résultats suggèrent une altération des processus liés à la familiarité et à la mémoire épisodique dans le contexte de présentation de stimuli déplaisants chez les patientes, alors que les processus pariétaux plus tardifs de récollection consciente telle que reflétée par la P300 et la LPC pariétales ne sont que peu ou pas atteints chez celles-ci. Donc, nos résultats mettent en lumière une différenciation des mécanismes du contrôle mnésique frontaux, relativement précoces, associés à la valence émotionnelle chez les patientes atteintes de schizophrénies.
Parties annexes
Remerciements
Ce travail a été soutenu en partie par le Conseil de recherche en sciences naturelles et génie du Canada (CRSNG), grâce à une subvention à la découverte attribuée à MEL et AM (# 355883) et grâce à un stage d’été du CRSNG octroyé à EG. Nous tenons à exprimer notre gratitude à notre technicienne Martine Germain, pour les enregistrements EEG, ainsi qu’à Mélissa Rinaldi, Jose Jimenez et Adam Mancini-Marie pour la coordination de différentes phases de la recherche, le dépistage clinique et la psychométrie. Les résultats de la présente recherche ont été présentés à l’occasion du 1er congrès sur les neurosciences affectives en santé mentale le 6 mai 2014. Nous remercions également toutes les participantes pour leur précieuse contribution à la présente étude.
Notes
-
[1]
Les composantes des potentiels évoqués cognitifs sont habituellement nommées par une lettre, soit le « P » ou le « N » représentent la polarité ou la valence électrique suivie d’un nombre, indiquant le temps d’apparition maximum de la composante, c’est-à-dire la latence en millisecondes après le début de la présentation du stimulus. Par exemple, la composante P300 reflète une déviation positive avec un pic autour de 300 ms suivant le stimulus. Il est aussi possible d’identifier une composante en fonction de sa distribution topographique sur le scalp. Ainsi, une composante P100 occipitale n’aurait pas la même signification qu’une P100 de distribution frontale. Une autre caractéristique intéressante des PÉC réside dans le fait qu’ils ne sont pas directement liés aux attributs physiques d’un stimulus (c.-à-d. exogènes), mais plutôt au traitement cognitif du stimulus (c.-à-d. endogènes).
-
[2]
Identification des images sélectionnées de la banque IAPS : valence plaisante/activation élevée : 1022 1030 1040 1070 1080 1090 1101 1110 1113 1114 1200 1205 1220 1240 1274 1300 1301 1310 1930 1931 3010 3022 3071 3250 3400 6020 6211 6530 6555 6836 6840 6910 6940 7560 8480 9102 9120 9140 9160 9230 9373 9452 9530 9560 9600 9621 9630 9635.1 9810 9920 | valence plaisante/activation faible : 1230 2055 2095 2110 2141 2276 2280 2312 2441 2491 2700 2722 2753 2795 2900.1 3300 4001 4003 4142 42104 2404 2794 2904 3024 4904 530 4561 4572 6561 7700 9045 9090 9110 9171 9190 9220 9290 9330 9331 9341 9342 9360 9390 9417 9440 9471 9472 9912 9913 | Valence déplaisante/activation élevée : 1303 1390 1650 2389 2616 2661 3005 4561 4598 4601 4606 4607 4608 4609 4651 4652 4664 4666 4672 4676 4677 4681 4687 4690 5450 5623 5626 5629 7402 7460 7501 7600 8021 8031 8034 8117 8130 8160 8161 8178 8179 8186 8192 8193 8200 8232 8251 8500 8503 9411 | Valence déplaisante/activation faible : 1313 1500 1600 1616 1661 1740 1810 1812 1942 2070 2170 2191 2260 2385 2370 2394 2442 2485 2487 2499 2500 2510 2515 2660 2749 4536 4625 4625 5760 5900 5991 7039 7095 7205 7207 7237 7284 7285 7286 7340 7493 7500 7503 7820 8032 8050 8220 8320 8600
Bibliographie
- Achim, A. M. & Lepage, M. (2005). Episodic memory-related activation in schizophrenia : meta-analysis. Br J Psychiatry, 187, 500-509. doi : 10.1192/bjp.187.6.500
- Addington, J. & Addington, D. (1998). Facial affect recognition and information processing in schizophrenia and bipolar disorder. Schizophr Res, 32(3), 171-181. doi : S0920996498000425 [pii]
- Addis, D. R., Leclerc, C. M., Muscatell, K. A. & Kensinger, E. A. (2010). There are age-related changes in neural connectivity during the encoding of positive, but not negative, information. Cortex, 46(4), 425-433. doi : 10.1016/j.cortex.2009.04.011
- Alain, C., Bernstein, L. J., Cortese, F., Yu, H. & Zipursky, R. B. (2002). Deficits in automatically detecting changes in conjunction of auditory features in patients with schizophrenia. Psychophysiology, 39(5), 599-606. doi : 10.1017.S0048577202394101
- American Psychiatric Association. (2000). Diagnostic and statistical manual of mental disorders, Fourth Edition. Text-revised. Washington, DC : auteur.
- An, S. K., Lee, S. J., Lee, C. H., Cho, H. S., Lee, P. G., Lee, C. I.,… Namkoong, K. (2003). Reduced P3 amplitudes by negative facial emotional photographs in schizophrenia. Schizophrenia research, 64(2-3), 125-135. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/14613677
- Arboleda-Florez. (1997). Consent in Psychiatry. http://cpa.medical.org/cpa/public2/papers/position.papers/consent.html
- Bar, M. (2003). A cortical mechanism for triggering top-down facilitation in visual object recognition. Journal of cognitive neuroscience, 15(4), 600-609. doi : 10.1162/089892903321662976
- Bar, M., Kassam, K. S., Ghuman, A. S., Boshyan, J., Schmid, A. M., Dale, A. M.,… Halgren, E. (2006). Top-down facilitation of visual recognition. Proceedings of the National Academy of Sciences of the United States of America, 103(2), 449-454. doi : 10.1073/pnas.0507062103
- Benedetti, F., Radaelli, D., Poletti, S., Falini, A., Cavallaro, R., Dallaspezia, S., ... Smeraldi, E. (2011). Emotional reactivity in chronic schizophrenia : structural and functional brain correlates and the influence of adverse childhood experiences. Psychol Med, 41(3), 509-519. doi : 10.1017/S0033291710001108
- Blanchard, J. J. & Cohen, A. S. (2006). The structure of negative symptoms within schizophrenia : implications for assessment. Schizophr Bull, 32(2), 238-245. doi : sbj013 [pii]10.1093/schbul/sbj013
- Canada, H. R. a. S. D. (2001). The National Occupational Classification and the Career Handbook. http://www5.hrsdc.gc.ca/noc/English/CH/2001/IntroductionNOCandCH.aspx
- Carretie, L., Hinojosa, J. A., Martin-Loeches, M., Mercado, F. & Tapia, M. (2004). Automatic attention to emotional stimuli : neural correlates. Hum Brain Mapp, 22(4), 290-299. doi : 10.1002/hbm.20037
- Carretie, L., Mercado, F., Hinojosa, J. A., Martin-Loeches, M. & Sotillo, M. (2004). Valence-related vigilance biases in anxiety studied through event-related potentials. J Affect Disord, 78(2), 119-130. doi : S0165032702002422 [pii]
- Champagne, J., Lakis, N., Bourque, J., Stip, E., Lipp, O. & Mendrek, A. (2012). Progesterone and Cerebral Function during Emotion Processing in Men and Women with Schizophrenia. Schizophr Res Treatment, 2012, 917901. doi : 10.1155/2012/917901
- Champagne, J., Mendrek, A., Germain, M., Hot, P. & Lavoie, M. E. (2014). Event-related brain potentials to emotional images and gonadal steroid hormone levels in patients with schizophrenia and paired controls. Front Psychol, 5, 543. doi : 10.3389/fpsyg.2014.00543
- Cohen, A. S. & Minor, K. S. (2010). Emotional experience in patients with schizophrenia revisited : meta-analysis of laboratory studies. Schizophr Bull, 36(1), 143-150. doi : sbn061 [pii]10.1093/schbul/sbn061
- Comerchero, M. D. & Polich, J. (1998). P3a, perceptual distinctiveness, and stimulus modality. Brain research. Cognitive brain research, 7(1), 41-48. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9714727
- Comerchero, M. D. & Polich, J. (1999). P3a and P3b from typical auditory and visual stimuli. Clinical neurophysiology : official journal of the International Federation of Clinical Neurophysiology, 110(1), 24-30. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10348317
- Curran, T. (1999). The electrophysiology of incidental and intentional retrieval : ERP old/new effects in lexical decision and recognition memory. Neuropsychologia, 37(7), 771-785. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10408645
- Curran, T. & Cleary, A. M. (2003). Using ERPs to dissociate recollection from familiarity in picture recognition. Brain Res Cogn Brain Res, 15(2), 191-205. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/12429370
- Curran, T. & Dien, J. (2003). Differentiating amodal familiarity from modality-specific memory processes : an ERP study. Psychophysiology, 40(6), 979-988. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/14986851
- Cuthbert, B. N., Schupp, H. T., Bradley, M. M., Birbaumer, N. & Lang, P. J. (2000). Brain potentials in affective picture processing : covariation with autonomic arousal and affective report. Biol Psychol, 52(2), 95-111. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10699350
- Diedrich O., N. E., Maier S., Becker G., Bartussek D. (1997). A frontal positive slow wave in the ERP associated with emotional slides. Journal of Psychophysiology, 11, 71-84.
- Dolcos, F. & Cabeza, R. (2002). Event-related potentials of emotional memory : encoding pleasant, unpleasant, and neutral pictures. Cogn Affect Behav Neurosci, 2(3), 252-263. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=12775189
- Duncan-Johnson, C. C. & Donchin, E. (1982). The P300 component of the event-related brain potential as an index of information processing. Biological psychology, 14(1-2), 1-52. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/6809064
- Egan, M. F., Duncan, C. C., Suddath, R. L., Kirch, D. G., Mirsky, A. F. & Wyatt, R. J. (1994). Event-related potential abnormalities correlate with structural brain alterations and clinical features in patients with chronic schizophrenia. Schizophrenia research, 11(3), 259-271. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/7910755
- First, M. B., Spitzer, R. L., Gibbon, M. & Williams, J. (1996). Structured Clinical Interview for DSM-IV Axis I Disorders. Biometrics Research Department New York State Psychiatric Institute, New York.
- Fletcher, P. C. & Frith, C. D. (2009). Perceiving is believing : a Bayesian approach to explaining the positive symptoms of schizophrenia. Nat Rev Neurosci, 10(1), 48-58. doi : 10.1038/nrn2536
- Folsom, D. P., Hawthorne, W., Lindamer, L., Gilmer, T., Bailey, A., Golshan, S.,… Jeste, D. V. (2005). Prevalence and risk factors for homelessness and utilization of mental health services among 10,340 patients with serious mental illness in a large public mental health system. Am J Psychiatry, 162(2), 370-376. doi : 162/2/370 [pii]10.1176/appi.ajp.162.2.370
- Folstein, J. R. & van Petten, C. (2011). After the P3 : late executive processes in stimulus categorization. Psychophysiology, 48(6), 825-841. doi : 10.1111/j.1469-8986.2010.01146.x
- Foxe, J. J., Doniger, G. M. & Javitt, D. C. (2001). Early visual processing deficits in schizophrenia : impaired P1 generation revealed by high-density electrical mapping. Neuroreport, 12(17), 3815-3820. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/11726801
- Friedman, D. (1990). Cognitive event-related potential components during continuous recognition memory for pictures. Psychophysiology, 27(2), 136-148. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/2247545b
- Gaebel, W. & Wolwer, W. (1992). Facial expression and emotional face recognition in schizophrenia and depression. Eur Arch Psychiatry Clin Neurosci, 242(1), 46-52. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=1390955
- Glaser, E., Mendrek, A., Germain, M., Lakis, N. & Lavoie, M. E. (2012). Sex differences in memory of emotional images : A behavioral and electrophysiological investigation. Int J Psychophysiol, 85(1), 17-26. doi : 10.1016/j.ijpsycho.2012.01.007
- Green, M. F. (2006). Cognitive impairment and functional outcome in schizophrenia and bipolar disorder. J Clin Psychiatry, 67(10), e12. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17107235
- Häfner, H., Riecher-Rössler, A., an der Heiden, W., Maurer, K., Fätkenheuer, B. & Löffler, W. (1993). Generating and testing a causal explanation of the gender difference in age at first onset of schizophrenia. Psychological Medicine, 23, 925-940.
- Halgren, E., Baudena, P., Clarke, J. M., Heit, G., Marinkovic, K., Devaux, B., . . . Biraben, A. (1995). Intracerebral potentials to rare target and distractor auditory and visual stimuli. II. Medial, lateral and posterior temporal lobe. Electroencephalogr Clin Neurophysiol, 94(4), 229-250. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/7537196
- Herry, C., Ciocchi, S., Senn, V., Demmou, L., Muller, C. & Luthi, A. (2008). Switching on and off fear by distinct neuronal circuits. Nature, 454(7204), 600-606. doi : 10.1038/nature07166
- Hill, H. & Weisbrod, M. (1999). The relation between asymmetry and amplitude of the P300 field in schizophrenia. Clinical neurophysiology : official journal of the International Federation of Clinical Neurophysiology, 110(9), 1611-1617. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10479028
- Hintzman, D. L., Caulton, D. A. & Levitin, D. J. (1998). Retrieval dynamics in recognition and list discrimination : further evidence of separate processes of familiarity and recall. Mem Cognit, 26(3), 449-462. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9610117
- Hoekert, M., Kahn, R. S., Pijnenborg, M. & Aleman, A. (2007). Impaired recognition and expression of emotional prosody in schizophrenia : review and meta-analysis. Schizophr Res, 96(1-3), 135-145. doi : S0920-9964(07)00322-2 [pii]10.1016/j.schres.2007.07.023
- Horan, W. P., Blanchard, J. J., Clark, L. A. & Green, M. F. (2008). Affective traits in schizophrenia and schizotypy. Schizophr Bull, 34(5), 856-874. doi : sbn083 [pii]10.1093/schbul/sbn083
- Horan, W. P., Wynn, J. K., Kring, A. M., Simons, R. F. & Green, M. F. (2010). Electrophysiological correlates of emotional responding in schizophrenia. J Abnorm Psychol, 119(1), 18-30. doi : 10.1037/a0017510
- Iguchi, Y. & Hashimoto, I. (2000). Sequential information processing during a mental arithmetic is reflected in the time course of event-related brain potentials. Clin Neurophysiol, 111(2), 204-213. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10680555
- Jeon, Y. W. & Polich, J. (2003). Meta-analysis of P300 and schizophrenia : patients, paradigms, and practical implications. Psychophysiology, 40(5), 684-701. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/14696723
- Jin, J. & Maren, S. (2015). Prefrontal-Hippocampal Interactions in Memory and Emotion. Front Syst Neurosci, 9, 170. doi : 10.3389/fnsys.2015.00170
- Kalisch, R., Korenfeld, E., Stephan, K. E., Weiskopf, N., Seymour, B. & Dolan, R. J. (2006). Context-dependent human extinction memory is mediated by a ventromedial prefrontal and hippocampal network. J Neurosci, 26(37), 9503-9511. doi : 10.1523/JNEUROSCI.2021-06.2006
- Kay, S. R., Fiszbein, A. & Opler, L. A. (1987). The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull, 13(2), 261-276. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=3616518
- Kim, A. S., Vallesi, A., Picton, T. W. & Tulving, E. (2009). Cognitive association formation in episodic memory : evidence from event-related potentials. Neuropsychologia, 47(14), 3162-3173. doi : 10.1016/j.neuropsychologia.2009.07.015
- Kim, M. A., Tura, E., Potkin, S. G., Fallon, J. H., Manoach, D. S., Calhoun, V. D.,… Turner, J. A. (2010). Working memory circuitry in schizophrenia shows widespread cortical inefficiency and compensation. Schizophr Res, 117(1), 42-51. doi : 10.1016/j.schres.2009.12.014
- Kissler, J., Herbert, C., Winkler, I. & Junghofer, M. (2009). Emotion and attention in visual word processing : an ERP study. Biol Psychol, 80(1), 75-83. doi : 10.1016/j.biopsycho.2008.03.004
- Knapska, E., Macias, M., Mikosz, M., Nowak, A., Owczarek, D., Wawrzyniak, M.,… Kaczmarek, L. (2012). Functional anatomy of neural circuits regulating fear and extinction. Proc Natl Acad Sci U S A, 109(42), 17093-17098. doi : 10.1073/pnas.1202087109
- Lakis, N., Jimenez, J. A., Mancini-Marie, A., Stip, E., Lavoie, M. E. & Mendrek, A. (2011). Neural correlates of emotional recognition memory in schizophrenia : effects of valence and arousal. Psychiatry Res, 194(3), 245-256. doi : 10.1016/j.pscychresns.2011.05.010
- Lang, P. J., Bradley, M. M. & Cuthbert, B. N. (2008). International Affective Picture System (IAPS) : Affective Ratings of Pictures and Instruction Manual. Gainesvillem FL. : University of Florida.
- Lavoie, M. E. & O’Connor, K. P. (2013). Effect of emotional valence on episodic memory stages as indexed by event-related potentials. World J Neurosci, 2013(3), 250-262. doi : 10.4236/wjns.2013.34034
- Lett, T. A., Voineskos, A. N., Kennedy, J. L., Levine, B. & Daskalakis, Z. J. (2014). Treating working memory deficits in schizophrenia : a review of the neurobiology. Biol Psychiatry, 75(5), 361-370. doi : 10.1016/j.biopsych.2013.07.026
- Luck, S. J. (2005). An Introduction of the Event-related Potential Technique. Cambridge.
- Mak, A. K., Hu, Z. G., Zhang, J. X., Xiao, Z. W. & Lee, T. M. (2009). Neural correlates of regulation of positive and negative emotions : an fmri study. Neuroscience letters, 457(2), 101-106. doi : 10.1016/j.neulet.2009.03.094
- Maratos, E. J., Allan, K. & Rugg, M. D. (2000). Recognition memory for emotionally negative and neutral words : an ERP study. Neuropsychologia, 38(11), 1452-1465. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10906371
- Meyer-Lindenberg, A. S., Olsen, R. K., Kohn, P. D., Brown, T., Egan, M. F., Weinberger, D. R. & Berman, K. F. (2005). Regionally specific disturbance of dorsolateral prefrontal-hippocampal functional connectivity in schizophrenia. Arch Gen Psychiatry, 62(4), 379-386. doi : 10.1001/archpsyc.62.4.379
- O’Donnell, B. F., Hokama, H., McCarley, R. W., Smith, R. S., Salisbury, D. F., Mondrow, E.,… Shenton, M. E. (1994). Auditory ERPs to non-target stimuli in schizophrenia : relationship to probability, task-demands, and target ERPs. International journal of psychophysiology : official journal of the International Organization of Psychophysiology, 17(3), 219-231. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/7806466
- O’Donnell, B. F., Shenton, M. E., McCarley, R. W., Faux, S. F., Smith, R. S., Salisbury, D. F.,… Jolesz, F. A. (1993). The auditory N2 component in schizophrenia : relationship to MRI temporal lobe gray matter and to other ERP abnormalities. Biological psychiatry, 34(1-2), 26-40. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8373937
- Ochsner, K. N., Bunge, S. A., Gross, J. J. & Gabrieli, J. D. (2002). Rethinking feelings : an FMRI study of the cognitive regulation of emotion. Journal of cognitive neuroscience, 14(8), 1215-1229. doi : 10.1162/089892902760807212
- Oldfield, R. C. (1971). The assessment and analysis of handedness : the Edinburgh inventory. Neuropsychologia, 9(1), 97-113. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=5146491
- Olofsson, J. K., Nordin, S., Sequeira, H. & Polich, J. (2008). Affective picture processing : an integrative review of ERP findings. Biol Psychol, 77(3), 247-265. doi : S0301-0511(07)00191-3 [pii]10.1016/j.biopsycho.2007.11.006
- Paller, K. A., Gonsalves, B., Grabowecky, M., Bozic, V. S. & Yamada, S. (2000). Electrophysiological correlates of recollecting faces of known and unknown individuals. Neuroimage, 11(2), 98-110. doi : 10.1006/nimg.1999.0521
- Pickering, E. C. & Schweinberger, S. R. (2003). N200, N250r, and N400 event-related brain potentials reveal three loci of repetition priming for familiar names. Journal of experimental psychology. Learning, memory, and cognition, 29(6), 1298-1311. doi : 10.1037/0278-7393.29.6.1298
- Pivik, R. T., Broughton, R. J., Coppola, R., Davidson, R. J., Fox, N. & Nuwer, M. R. (1993). Guidelines for the recording and quantitative analysis of electroencephalographic activity in research contexts. Psychophysiology, 30(6), 547-558. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8248447
- Polich, J. (2007). Updating P300 : an integrative theory of P3a and P3b. Clin Neurophysiol, 118(10), 2128-2148. doi : S1388-2457(07)00189-7 [pii]10.1016/j.clinph.2007.04.019
- Preston, A. R. & Eichenbaum, H. (2013). Interplay of hippocampus and prefrontal cortex in memory. Curr Biol, 23(17), R764-773. doi : 10.1016/j.cub.2013.05.041
- Ragland, J. D., Laird, A. R., Ranganath, C., Blumenfeld, R. S., Gonzales, S. M. & Glahn, D. C. (2009). Prefrontal activation deficits during episodic memory in schizophrenia. Am J Psychiatry, 166(8), 863-874. doi : 10.1176/appi.ajp.2009.08091307
- Reichenberg, A. & Harvey, P. D. (2007). Neuropsychological impairments in schizophrenia : Integration of performance-based and brain imaging findings. Psychol Bull, 133(5), 833-858. doi : 10.1037/0033-2909.133.5.833
- Roth, W. T. & Cannon, E. H. (1972). Some features of the auditory evoked response in schizophrenics. Archives of general psychiatry, 27(4), 466-471. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/5072716
- Ruchkin, D. S., Johnson, R., Jr., Canoune, H. & Ritter, W. (1991). Event-related potentials during arithmetic and mental rotation. Electroencephalogr Clin Neurophysiol, 79(6), 473-487. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/1721575
- Ruchkin, D. S., Johnson, R., Jr., Mahaffey, D. & Sutton, S. (1988). Toward a functional categorization of slow waves. Psychophysiology, 25(3), 339-353. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/3406333
- Rugg, M. D., Roberts, R. C., Potter, D. D., Pickles, C. D. & Nagy, M. E. (1991). Event-related potentials related to recognition memory. Effects of unilateral temporal lobectomy and temporal lobe epilepsy. Brain, 114 ( Pt 5), 2313-2332. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/1933247
- Rugg, M. D., Walla, P., Schloerscheidt, A. M., Fletcher, P. C., Frith, C. D. & Dolan, R. J. (1998). Neural correlates of depth of processing effects on recollection : evidence from brain potentials and positron emission tomography. Exp Brain Res, 123(1-2), 18-23. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9835388
- Salisbury, D. F., O’Donnell, B. F., McCarley, R. W., Shenton, M. E. & Benavage, A. (1994). The N2 event-related potential reflects attention deficit in schizophrenia. Biological psychology, 39(1), 1-13. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/7880944
- Schneider, F., Weiss, U., Kessler, C., Salloum, J. B., Posse, S., Grodd, W. & Muller-Gartner, H. W. (1998). Differential amygdala activation in schizophrenia during sadness. Schizophr Res, 34(3), 133-142. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9850979
- Schupp, H. T., Junghofer, M., Weike, A. I. & Hamm, A. O. (2003). Attention and emotion : an ERP analysis of facilitated emotional stimulus processing. Neuroreport, 14(8), 1107-1110. doi : 10.1097/01.wnr.0000075416.59944.49
- Schupp, H. T., Junghofer, M., Weike, A. I. & Hamm, A. O. (2004). The selective processing of briefly presented affective pictures : an ERP analysis. Psychophysiology, 41(3), 441-449. doi : 10.1111/j.1469-8986.2004.00174.xPSYP174 [pii]
- Sigurdsson, T., Stark, K. L., Karayiorgou, M., Gogos, J. A. & Gordon, J. A. (2010). Impaired hippocampal-prefrontal synchrony in a genetic mouse model of schizophrenia. Nature, 464(7289), 763-767. doi : 10.1038/nature08855
- Spaniol, J., Davidson, P. S., Kim, A. S., Han, H., Moscovitch, M. & Grady, C. L. (2009). Event-related fMRI studies of episodic encoding and retrieval : meta-analyses using activation likelihood estimation. Neuropsychologia, 47(8-9), 1765-1779. doi : 10.1016/j.neuropsychologia.2009.02.028
- Stefansson, S. B. & Jonsdottir, T. J. (1996). Auditory event-related potentials, auditory digit span, and clinical symptoms in chronic schizophrenic men on neuroleptic medication. Biological psychiatry, 40(1), 19-27. doi : 10.1016/0006-3223(95)00351-7
- Stewart, J. L., Silton, R. L., Sass, S. M., Fisher, J. E., Edgar, J. C., Heller, W. & Miller, G. A. (2010). Attentional bias to negative emotion as a function of approach and withdrawal anger styles : an ERP investigation. Int J Psychophysiol, 76(1), 9-18. doi : 10.1016/j.ijpsycho.2010.01.008
- Strauss, G. P., Kappenman, E. S., Culbreth, A. J., Catalano, L. T., Lee, B. G. & Gold, J. M. (2013). Emotion regulation abnormalities in schizophrenia : cognitive change strategies fail to decrease the neural response to unpleasant stimuli. Schizophr Bull, 39(4), 872-883. doi : 10.1093/schbul/sbs186
- Takahashi, H., Koeda, M., Oda, K., Matsuda, T., Matsushima, E., Matsuura, M.,… Okubo, Y. (2004). An fMRI study of differential neural response to affective pictures in schizophrenia. Neuroimage, 22(3), 1247-1254. doi : 10.1016/j.neuroimage.2004.03.028
- Taylor, S. F., Kang, J., Brege, I. S., Tso, I. F., Hosanagar, A. & Johnson, T. D. (2012). Meta-analysis of functional neuroimaging studies of emotion perception and experience in schizophrenia. Biol Psychiatry, 71(2), 136-145. doi : S0006-3223(11)00901-2 [pii]10.1016/j.biopsych.2011.09.007
- Tsai, P. C., McDowd, J., Tang, T. C. & Su, C. Y. (2012). Processing speed mediates gender differences in memory in schizophrenia. Clin Neuropsychol, 26(4), 626-640. doi : 10.1080/13854046.2012.678887
- van der Werf, M., Kohler, S., Verkaaik, M., Verhey, F., van Os, J. & Investigators, G. (2012). Cognitive functioning and age at onset in non-affective psychotic disorder. Acta Psychiatr Scand, 126(4), 274-281. doi : 10.1111/j.1600-0447.2012.01873.x
- Van Strien, J. W., Langeslag, S. J., Strekalova, N. J., Gootjes, L. & Franken, I. H. (2009). Valence interacts with the early ERP old/new effect and arousal with the sustained ERP old/new effect for affective pictures. Brain Res, 1251, 223-235. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=19063866
- Verleger, R. (2008). P3b : towards some decision about memory. Clinical neurophysiology : official journal of the International Federation of Clinical Neurophysiology, 119(4), 968-970. doi : 10.1016/j.clinph.2007.11.175
- Whyte, M. C., Whalley, H. C., Simonotto, E., Flett, S., Shillcock, R., Marshall, I., ... Lawrie, S. M. (2006). Event-related fMRI of word classification and successful word recognition in subjects at genetically enhanced risk of schizophrenia. Psychol Med, 36(10), 1427-1439. doi : 10.1017/S0033291706008178
- Wilding, E. L. & Rugg, M. D. (1996). An event-related potential study of recognition memory with and without retrieval of source. Brain, 119 ( Pt 3), 889-905. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8673500
- Windmann, S. & Kutas, M. (2001). Electrophysiological correlates of emotion-induced recognition bias. J Cogn Neurosci, 13(5), 577-592. doi : 10.1162/089892901750363172
- Woods, S. W. (2003). Chlorpromazine equivalent doses for the newer atypical antipsychotics. J Clin Psychiatry, 64(6), 663-667. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=12823080
- Zhou, Y., Shu, N., Liu, Y., Song, M., Hao, Y., Liu, H.,… Jiang, T. (2008). Altered resting-state functional connectivity and anatomical connectivity of hippocampus in schizophrenia. Schizophr Res, 100(1-3), 120-132. doi : 10.1016/j.schres.2007.11.039
Liste des figures
Figure 1
Amplitude de la N200 frontale en réponse aux conditions de mémoire épisodique connu (cercle noir) et inconnu (triangle blanc) en fonction des groupes et des conditions de valence émotionnelle. En dépit d’une N200 frontale généralement réduite, les patientes démontrent une augmentation de l’effet de mémoire N200 dans l’hémisphère droit par rapport à l’hémisphère gauche, en réponse aux stimuli déplaisants. Pour ce qui est du groupe contrôle, il y a aussi une augmentation de cet effet de mémoire dans l’hémisphère droit, mais cette fois, en réponse aux stimuli plaisants.
Figure 2
Amplitude de la P300 frontale en réponse aux conditions de mémoire épisodique connu (cercle noir) et inconnu (triangle blanc) en fonction des groupes et des conditions de valence émotionnelle. Les patientes démontrent une augmentation de l’effet de mémoire P300 dans l’hémisphère droit, uniquement en réponse aux stimuli déplaisants. Le groupe contrôle démontre un plus grand effet de mémoire dans l’hémisphère droit en réponse aux stimuli plaisants.
Figure 3
Illustration des potentiels évoqués cognitifs frontaux de l’hémisphère gauche et droit moyennés pour les images plaisantes (panneau A) et déplaisantes (panneau B). Les patientes schizophrènes dénotent une N200 réduite, alors que la P300 et la LPC sont généralement plus amples que chez les contrôles.
Figure 4
Illustration des potentiels évoqués cognitifs pariétaux de l’hémisphère gauche et droit moyennés pour les images plaisantes (panneau A) et déplaisantes (panneau B). L’effet de mémoire sur la N200 pariétale est généralement réduit chez les patientes schizophrènes en comparaison au groupe contrôle. Plus loin dans le décours temporel, le groupe de patientes dénote une amplitude de la composante P300 significativement plus importante, mais l’effet de mémoire épisodique est toutefois réduit dans ce groupe, et ce, dans toutes les conditions émotionnelles.
Liste des tableaux
Tableau 1
Comparaison des moyennes et écarts-types (parenthèses) des caractéristiques démographiques et cliniques
* ; p < ,05 ** p < ,005
WAIS : Weschler Adult Intelligence Scale ; PANSS : Positive and Negative Syndrome Scale, SSE= Statut socioéconomique ; NS : non significatif
Tableau 2
Comparaison des moyennes et écarts-types (parenthèses) du bilan hormonal
* p < ,05 ; ** p < ,005
Tableau 3
Comparaison des évaluations subjectives de la valence et de l’activation émotionnelle en fonction des types de réponses en mémoire (connus vs inconnus) dans les deux groupes, ainsi que pour la norme IAPS pour les femmes
Valence : Évaluation de la valence émotionnelles selon une échelle de 1 à 9 (1 = très déplaisant | 5 = neutre | 9 = très plaisants)
Activation : Évaluation de l’activation émotionnelle selon une échelle de 1 à 9 (1= activation très faible à 9 = activation très forte)
Tableau 4
Comparaison des temps de réaction en millisecondes (erreur-type) en réponse à la valence (plaisantes et déplaisantes) et à l’activation (élevées et faibles) émotionnelle en fonction des types de réponses en mémoire (connus vs inconnus) dans les deux groupes
Note : Pl = valence plaisantes ; Dpl = valence déplaisantes ; É = activations élevées ; F = activations faibles