
Résumés
Résumé
De manière générale, il apparaît que l’acte suicidaire est hautement imprévisible par les moyens scientifiques actuellement à notre disposition. Dans cet article, nous formulons l’hypothèse que la prédiction du suicide est compliquée parce qu’elle résulte de la prédiction d’un choix, lequel est lui-même imprévisible. Nous proposons un modèle du suicide selon l’apprentissage par renforcement. Dans ce modèle, nous intégrons d’une part les quatre principaux modulateurs ascendants (acétylcholine, noradrénaline, sérotonine, dopamine) avec leurs régions de projections et d’afférences respectives, et d’autre part, différentes observations d’imagerie cérébrales trouvées à ce jour dans le processus suicidaire.
Abstract
In general, it appears that the suicidal act is highly unpredictable with the current scientific means available. In this article, the author submits the hypothesis that predicting suicide is complex because it results in predicting a choice, in itself unpredictable. The article proposes a Reinforcement learning model-based analysis. In this model, we integrate on the one hand, four ascending modulatory neurotransmitter systems (acetylcholine, noradrenalin, serotonin, and dopamine) with their regions of respective projections and afferences, and on the other hand, various observations of brain imaging identified until now in the suicidal process.
Corps de l’article
Les prédictions d’événements inattendus sont toujours plus précises si on ne les a pas rédigées auparavant.
Carl Sagan, Contact
Prédire le suicide ?
Le suicide est un problème de santé publique majeur et une cause importante de décès chaque année. Environ 13,5 % de la population des USA a eu des idées suicidaires ou un désir d’en finir avec la vie à un moment donné, 3,9 % a eu un plan suicidaire, et 4,6 % a fait une tentative. La National Comorbidity Survey (Kessler et al., 1999) a montré que les probabilités cumulées étaient de 34 % pour la transition d’une idéation à un plan, de 72 % pour la transition d’un plan à une tentative, et de 26 % pour la transition d’une idéation sans plan à une tentative non planifiée. De plus, environ 90 % des tentatives non planifiées et 60 % des premières tentatives planifiées arrivaient dans la première année après la survenue d’idéations suicidaires (Kessler et al., 1999). Une autre étude récente sur 84 850 adultes de 17 pays a montré que dans tous les pays, 60 % des transitions des idéations aux plans et aux tentatives sont survenues dans la première année après le début des idéations suicidaires (Nock et al., 2008).
Ces chiffres indiquent qu’il serait important de pouvoir mesurer les idéations suicidaires pour prédire les tentatives de suicide. Cependant, malgré l’existence d’une vaste littérature sur les facteurs de risque des tentatives, les études prospectives n’ont pas réussi à prédire ne fut-ce qu’un suicide (Goldstein et al., 1991) (avec quelques exceptions pour deux études prospectives a long terme (Brown et al., 2000 ; Oquendo et al., 2004). Ainsi, la plupart des études n’ont pas réussi, ou ont réussi de manière limitée, à mettre en évidence un marqueur objectif pouvant prédire les tentatives de suicide ou un suicide accompli, que ce soit un marqueur psychologique (Wenzel et al., 2008) ou biologique (Mann et al., 2006).
Quelles sont les raisons de cette incapacité à prévoir le passage à l’acte ? Premièrement, il faut souligner que le suicide est un comportement avec un faible taux de base. Beaucoup de personnes peuvent éprouver que la vie ne vaut pas la peine d’être vécue, mais moins de personnes pensent à la mort, encore moins font une tentative de suicide et une toute petite partie seulement mourra de suicide. Ainsi, malgré que la plupart des gens ayant fait une tentative présentent des antécédents d’idéations suicidaires, il y a au total peu de personnes avec des idéations suicidaires qui feront une tentative. Deuxièmement, les patients dénient fréquemment leurs idéations suicidaires dans les dernières communications qu’ils font avant de se donner la mort (Busch et al., 2003). Ceci pourrait expliquer de façon pertinente le nombre élevé de suicides constaté à la sortie d’hospitalisation (Qin and Nordentoft, 2005), alors que le risque suicidaire évalué devrait être au plus bas à ce moment-là. Troisièmement, il s’agirait du peu de spécificité des marqueurs de prédiction clinique utilisés (Caspi et al., 2003) et du manque de reproductibilité (Risch et al., 2009). Quatrièmement, la raison serait le manque de précision des échelles cliniques utilisées, qui pourraient ne pas cerner adéquatement le processus de suicide en cours (Desseilles et al., 2011).
Une approche complémentaire à la mise en évidence de marqueurs psychologiques et biologiques est l’identification de marqueurs comportementaux. Au moyen de l’Implicit Association Task (i.e. tâche mettant en évidence les associations implicites entre soi-même et des mots ou des images), Nock et al. (2010) ont montré qu’une tâche comportementale était potentiellement intéressante dans la prédiction du risque de passage à l’acte (Nock et Banaji 2007 ; Nock et al., 2010). L’association implicite de « soi » avec les thèmes « mort/suicide » était associée à une augmentation de probabilité d’environ six fois de faire une tentative dans les six mois suivants. Cela surpassait la validité prédictive de facteurs de risque connus (par exemple, la dépression et les antécédents de tentative de suicide) mais aussi les prédictions des patients et des cliniciens (Nock et al., 2010).
Ces constatations nous amènent à conclure que l’acte suicidaire est hautement imprévisible par les moyens scientifiques actuellement à notre disposition.
Dans cet article, nous formulons l’hypothèse que la prédiction du suicide pourrait être rendue compliquée parce qu’elle résulte de la prédiction d’un choix, lequel est lui-même imprévisible. Afin d’étayer cette hypothèse, nous allons d’abord voir que le suicide peut être compris comme un choix. Nous verrons ensuite comment les choix imprévisibles peuvent être étudiés et enfin, nous proposerons des pistes d’étude du suicide.
Le suicide comme choix
Revoyons d’abord quelques définitions. Le suicide est une mort causée de façon intentionnelle par soi-même. L’intentionnalité est ce qui différencie un décès par suicide d’une mort causée de façon accidentelle par l’individu lui-même. Les tentatives de suicide sont des actes intentionnels commis par une personne, dans le but de se tuer, même si ces actes n’aboutissent pas toujours à la mort. Les idées suicidaires désignent des idées sérieuses d’en finir avec la vie (Wenzel et al., 2009).
L’intentionnalité du suicide suggère que le suicide puisse être considéré comme une prise de décision. La prise de décision n’est en fait qu’une composante des habiletés de résolution de problèmes. Celles-ci ont été définies par D’Zurilla et Goldfried (1971) en 5 étapes : l’orientation générale, la définition et la formulation du problème, la génération d’alternatives, la prise de décision et la vérification. Dans le processus de résolution de problèmes, certaines pensées ou comportements seront donc choisis afin d’arriver à une solution. Chez les sujets suicidaires, les habiletés de résolution de problèmes et la prise de décision peuvent être évaluées en utilisant des tâches neuropsychologiques, comme l’Iowa Gambling Task ou le Means-Ends Problem-Solving Procedure (e.g., Oldershaw et al., 2009).
Comme nous le verrons plus bas, nous évaluons habituellement l’incertitude d’une récompense dans l’environnement d’une manière optimale et nous ajustons notre prise de décision en fonction de notre évaluation (Behrens et al., 2007).
Cependant, ce processus apparaît être différent chez les individus suicidaires. Ces derniers montrent des anomalies dans des domaines spécifiques nécessaires à la prise de décision, et ont donc des difficultés dans la résolution des problèmes. Ils génèrent peu de solutions alternatives à une situation particulière et ils cherchent des solutions rapides (i.e. avec des conséquences positives à court terme) sans tenir compte des conséquences potentiellement néfastes à long terme. Pour Chiles et Strosahl (2005), ce style de résolution de problèmes mène l’individu suicidaire à considérer le comportement suicidaire comme une stratégie efficace de résolution de problèmes.
En plus de la faible capacité à générer des alternatives, un biais de traitement de l’information est observé chez les patients qui présentent des idées suicidaires (Wenzel et al., 2009). Ces patients montrent des biais attentionnels congruents avec le contenu des pensées liées au suicide. Ce biais rétrécit dès lors leur champ attentionnel en favorisant l’attention vers des stimuli ayant trait directement aux contenus suicidaires et en les éloignant ainsi des autres alternatives. Ce phénomène est appelé la fixation attentionnelle (Williams et Broadbent, 1986 ; Becker et al., 1999).
Les sujets suicidaires sont caractérisés également par un biais mnésique de surgénéralisation. Ce biais les empêche d’accéder à des informations spécifiques de leur passé qui pourraient les aider à prendre des décisions dans le présent et à résoudre leurs problèmes (Williams et al., 2006 ; Behrens et al., 2007). C’est ce que Shneidman (1985) appelle la constriction cognitive.
La combinaison de ces biais attentionnels et mnésiques conduit à des anomalies de flexibilité cognitive et à une dissociation des processus réflexifs de haut niveau. Ces deux anomalies interfèrent avec la prise de décision et donc avec la résolution de problèmes (figure 1).
Figure 1
Déficits pour la résolution de problèmes

Les stress de notre vie et les tracas quotidiens agissent sur nos processus cognitifs et en particulier sur les cognitions en rapport avec le suicide, sous forme de pensées ou d’interprétations. Ces modifications cognitives influent sur le traitement de l’information, premièrement à court terme, via des biais attentionnels qui amènent le sujet à diriger son attention vers des stimuli négatifs, deuxièmement à long terme, via des biais mnésiques résultant d’une mémoire surgénéralisée. En particulier, la fixation attentionnelle et la constriction cognitive entraînent des déficits pour la résolution de problèmes.
Nous avons vu plus haut que le suicide était difficilement prévisible par les moyens scientifiques actuels. Le suicide est une décision prise à un certain moment donné dans le cadre d’une résolution de problèmes. Les patients suicidaires ont un mode particulier de prise de décision et de résolution de problèmes qui les conduit à choisir une solution rapide, sans tenir compte des conséquences négatives à long terme. Si le suicide est l’aboutissement d’une décision, celle-ci peut être prévisible lorsque la décision est prise de manière posée et résulte du fruit d’une réflexion calme et décidée. La prise de décision peut également être incertaine et imprévisible tant pour celui qui commet potentiellement l’acte suicidaire qu’a fortiori pour son environnement. Il y a bien sûr une multitude de cas de figure entre ces deux extrêmes.
Que ce soit au sujet du suicide ou de questions plus générales, nos choix sont guidés par nos expériences antérieures et par les imprévus de l’environnement. Nous allons voir maintenant que la nature imprévisible du choix peut toutefois être étudiée. Nous verrons aussi que l’imprévisibilité des choix a des corrélats cérébraux qui peuvent s’intégrer dans un réseau d’aires cérébrales et de neurotransmetteurs interagissant entre eux.
L’imprévisible nature du choix
Les humains semblent intégrer les informations visuelles et relevant du toucher d’une manière statistiquement optimale (Ernst et Banks, 2002). Il en serait de même pour l’apprentissage sensorimoteur (Kording et Wolpert, 2004). Néanmoins, nous aimons à penser que nous ne sommes pas faits que de perception, de motricité et d’arcs réflexes moteurs non régis par une volonté propre. Nous aimons à penser également qu’un choix libre et volontaire n’est pas simplement régi par des règles mathématiques, fussent-elles simples ou compliquées. En accord avec ces pensées issues de la psychologie populaire, le choix volontaire qui peut être considéré comme un processus de haut niveau hiérarchique (e.g. par rapport à la perception de stimuli visuels simples qui peut être considérée comme un processus de bas niveau hiérarchique) semble bien avoir une nature imprévisible (Kahneman et Tversky, 2000).
Cependant, des travaux récents mettent à l’épreuve cette perspective d’imprévisibilité du choix volontaire, et proposent qu’une estimation de certaines caractéristiques de l’environnement puisse influencer la manière dont nous prenons nos décisions volontaires (Behrens et al., 2007).
Nous sommes forcés de prendre des décisions tous les jours et certaines de nos décisions entraînent des résultats positifs. D’autres engendrent des conséquences négatives et d’autres encore semblent ne pas avoir de conséquences. Au fur et à mesure de notre évolution, nos décisions volontaires sont influencées par les résultats des décisions volontaires que nous avons prises dans le passé (Montague et al., 1995 ; Bayer et Glimcher, 2005 ; Samejima et al., 2005 ; Daw et al., 2006). Il semble ainsi logique qu’une meilleure compréhension des associations entre les décisions volontaires (les actions) et leurs résultats permettrait de modéliser le choix volontaire. C’est ce que propose la Théorie de l’apprentissage (voir figure 2).
Figure 2
Théorie de l’apprentissage

Selon le modèle d’apprentissage par renforcement (Sutton et Barto, 1998), lorsque nous recevons de nouvelles informations, nous mettons à jour nos croyances à propos de l’environnement, en proportion de l’erreur de prédiction (c’est-à-dire la différence entre l’issue escomptée et l’issue actuelle d’une action, le δ sur la figure). Ce doit être multiplié par un facteur (α) appelé le taux d’apprentissage, afin de déterminer le degré par lequel la valeur de l’action est mise à jour.
Les apports bayésiens suggèrent que le facteur α devrait dépendre du niveau actuel d’incertitude dans l’estimation de la valeur de l’action. Cette incertitude est déterminée par les statistiques de récompenses de l’environnement (par exemple Yu et Dayan, 2005).
Ainsi, les événements du passé ne devraient pas avoir la même influence sur nos décisions du moment. Nous proposons les deux exemples suivants pour illustrer notre propos. Un joueur d’échec puise dans son savoir pour bouger un pion en tenant compte des possibilités de mouvements de son adversaire et des multiples combinaisons. Ces dernières débouchent immanquablement sur d’autres possibilités ou d’autres contraintes qui, en fonction de l’habilité du joueur et de son adversaire, peuvent conduire vers une victoire ou un échec. Ainsi, aux échecs, la temporalité passée, présente et future est omniprésente, et le poids du passé (des résultats obtenus à la suite des coups passés) est prépondérant. Dans cet environnement lentement changeant, les événements du passé prennent un poids plus important dans la réactualisation des résultats attendus suite à une décision volontaire. Par contre, un joueur d’escrime doit tenir compte d’environnements beaucoup plus proximaux, en plus des événements distaux qui constituent son background. Dans cet environnement rapidement changeant, où l’adversaire peut à tout moment surprendre, il doit pouvoir rapidement mettre à jour ses croyances et ses pensées par rapport aux effets qu’ont ses actions.
Ces deux exemples nous montrent qu’en fonction de l’instabilité et de l’imprévisibilité de l’environnement, l’impact que les résultats de nos actions auront sur la mise à jour de nos croyances par rapport à cet environnement sera différent. Ainsi, on pourrait s’attendre à des décisions prises plus rapidement dans un environnement instable et imprévisible. On pourrait également supposer une mise à jour plus rapide mais aussi un plus grand stress (cognitif, affectif, exécutif), vu le recrutement plus important des ressources aux seules fins de la décision du choix d’action.
En termes de théorie de l’apprentissage, lorsque l’expérience actuelle prédit davantage le futur que l’expérience ancienne (e.g. joueur d’escrime), le taux d’apprentissage α devrait être grand (pour permettre au joueur d’escrime d’adapter rapidement son jeu pour ne pas être touché). Par contre, quand c’est l’expérience ancienne qui prédit plus le futur que l’expérience actuelle (e.g. jeu d’échec), le taux d’apprentissage α est petit (e.g. pour permettre au joueur d’accorder une importance aux données plus anciennes). Ainsi, le taux d’apprentissage α devrait maximiser la possibilité de prédire l’issue des actions. Prévoir l’issue d’un comportement, c’est bien le rôle de l’apprentissage !
Corrélats cérébraux de l’imprévisibilité et de la prise de décision
Y a-t-il une région cérébrale ou un circuit cérébral spécialisé dans l’imprévisibilité et l’instabilité des actions ? Y-a-t-il un neuromodulateur ou un groupe de modulateurs s’occupant de cette imprévisibilité ?
Les modèles théoriques distinguent d’abord l’imprévisibilité prévue de l’association entre un stimulus et son issue, et ensuite l’incertitude imprévue causée par le changement de ces imprévus (Yu et Dayan, 2005). Ces deux formes d’imprévisibilité se combineraient pour orienter nos comportements et donc nos choix.
L’incertitude prévue serait représentée dans les noyaux cholinergiques (Yu et Dayan, 2005). L’incertitude imprévue serait déterminée par la noradrénaline (Yu et Dayan, 2005) et ses interactions avec la partie antérieure du cortex cingulaire (ACC) (Aston-Jones et Cohen, 2005). De manière intéressante, une récente étude a montré que les humains estimaient l’imprévisibilité d’une manière optimale et ajustaient leurs prises de décision en accord avec cette imprévisibilité (Behrens et al., 2007). De plus, l’activité dans l’ACC corrélait avec le taux d’apprentissage et était en lien avec l’imprévisibilité de l’environnement estimée (Behrens et al., 2007). Ces résultats suggèrent que l’ACC et la noradrénaline pourraient être de bons candidats en tant que corrélats cérébraux de choix imprévisibles (comme celui de se suicider).
L’ACC est une région cérébrale qui fait partie d’un réseau neuronal impliqué dans la représentation et la mise à jour de la valeur de nos décisions (McCoy et al., 2003 ; Bayer et Glimcher,, 2005; Samejima et al., 2005). D’une part, la prédiction de signaux d’erreur est en lien avec l’activité de régions dopaminergiques (Waelti et al., 2001) et du striatum ventral (O’Doherty et al., 2004 ; Montague et al., 2006). D’autre part, la valeur de l’action serait en lien avec l’activité dans le putamen (Haruno et al., 2004 ; Tanaka et al., 2004). Ainsi, les projections de l’ACC vers le striatum ventral (Kunishio et Haber, 1994) pourraient permettre au taux d’apprentissage α de moduler l’influence de l’erreur de prédiction actuelle sur l’estimation de la valeur de l’action (Rescorla et Wagner, 1972).
Un patient avec des idées suicidaires pourrait être confronté à un choix imprévisible impliquant l’ACC et la noradrénaline. Ce patient pourrait réévaluer la valeur de son action de manière répétée et ainsi solliciter tout particulièrement l’ACC, le striatum ventral et le putamen. De manière congruente à cette hypothèse, l’ACC (Oquendo et al., 2003 ; Monkul et al., 2007 ; Jollant et al., 2008), le striatum ventral et le putamen (Dombrovski et al., 2011 ; Marchand et al., 2012) ont été impliqués dans la physiopathologie du processus suicidaire.
Substrats neurobiologiques du processus suicidaire
Nous venons de voir que l’incertitude imprévue impliquait la noradrénaline. Ce neuromodulateur est libéré au niveau du tronc cérébral dans une région appelée le locus coeruleus (LC). Un modèle de fonctionnement du système LC-noradrénaline a été proposé par Aston-Jones et ses collaborateurs. Dans ce modèle, il propose que la balance entre l’exploitation (i.e. l’utilisation au mieux de ses performances pour la réalisation d’une tâche) et l’exploration (i.e. l’exploration de son environnement pour trouver des alternatives) se trouvent sur une courbe en U inversé (Aston-Jones et al., 1996 ; Aston-Jones et Cohen, 2005). Selon cette théorie, une augmentation de la noradrénaline, du stress ou de l’éveil (i.e. arousal) à un niveau modéré faciliterait les comportements d’exploitation. Cependant, à un taux très élevé de noradrénaline, les changements (« switch ») attentionnels seraient plus fréquents et la concentration décroîtrait et détériorerait les performances à une tâche (Aston-Jones et Cohen, 2005). Ce genre de situation comportant un stress élevé conduirait à une prise de choix impulsive, associée à une récompense à court terme (Luksys et al., 2009). Chez l’animal, de très hauts taux de noradrénaline ont été associés à des observations comportementales similaires, c’est-à-dire une attention rapidement changeante et beaucoup de changements de stratégies (Aston-Jones et al., 2000 ; Aston-Jones et Cohen, 2005). Ces observations de changements rapides de stratégies sont congruentes avec l’incertitude imprévue.
Selon le modèle stress-diathèse du suicide de J. John Mann, une faible quantité de sérotonine conduit à une impulsivité accrue tandis qu’une faible quantité de noradrénaline mène au désespoir ou au pessimisme, ces deux anomalies convergeant vers l’acte suicidaire (Mann, 2003).
Par ailleurs, Mann suggère que l’hyperactivité noradrénergique corticale rencontrée chez les individus ayant fait une tentative de suicide pourrait conduire à la déplétion subséquente de la noradrénaline dans les populations de neurones qui s’avèrent être en sous-nombre chez les victimes de suicide (Mann, 2003, p. 822). De plus, cet auteur propose une hypothèse pour expliquer la régulation à la hausse de la tyrosine hydroxylase (i.e., l’enzyme limitant la biosynthèse de la noradrénaline) observée dans le LC des victimes de suicide. Cette régulation pourrait constituer une réponse au relargage excessif de noradrénaline généré par le stress du suicide imminent (Mann, 2003, p. 822). Cette suggestion est congruente avec l’augmentation observée de la sécrétion de noradrénaline peu de temps après une tentative de suicide (Mancini et Brown, 1992). Cette augmentation de noradrénaline suggère une augmentation d’activité dans le LC et a été liée à des choix impulsifs (Eckhoff et al.,, 2009) ainsi qu’à des changements comportementaux rapides en réponse à des challenges cognitifs, par la facilitation de la réorganisation dynamique des réseaux corticaux (Bouret et Sara, 2005). Cette hypothèse s’appuie aussi sur l’observation qu’une prise brève d’antidépresseur chez des jeunes rats augmente les décharges des neurones dans le LC, et est associé avec une péjoration de la dépression (West et al., 2010), suggérant par là que l’augmentation d’activité du LC puisse être associée aux effets secondaires du traitement antidépresseur sur l’humeur particulièrement chez les jeunes.
Enfin, l’investigation des marqueurs génétiques prédisant l’augmentation des idéations suicidaires suggère que chez les hommes, les variations génétiques dans la signalisation noradrénergique pourraient interagir avec les antidépresseurs inhibiteurs de la recapture de la noradrénaline, et ainsi contribuer à l’augmentation de la suicidalité observée avec la nortriptyline (Perroud et al., 2009).
Toutes ces observations suggèrent que le système LC-noradrénaline pourrait être impliqué dans l’incertitude imprévue, la prise de décision rapide et des changements comportementaux rapides tels que ceux observés dans le processus suicidaire.
Le système LC-noradrénaline est un des nombreux noyaux neuromodulateurs du tronc cérébral qui se distribue très largement via des projections ascendantes dans le néocortex (Aston-Jones et Cohen, 2005). En particulier, le LC reçoit des informations à propos des récompenses et des coûts via des projections de deux régions du cortex frontal : le cortex orbitofrontal (COF) et l’ACC. Le COF a été associé à l’évaluation de la récompense et l’ACC à l’évaluation des coûts (Aston-Jones et Cohen, 2005). Ces deux structures frontales corticales sont importantes dans la balance entre l’exploitation et l’exploration. De manière congruente, des anomalies fronto-limbiques impliquant spécifiquement le COF ont été observées chez les patients suicidaires (Monkul et al., 2007 ; Jollant et al.,, 2008; Jollant et al.,, 2010).
Le LC envoie et reçoit des projections du cortex préfrontal, incluant l’ACC. Cette boucle entre le cortex préfrontal et le LC offre un mécanisme à travers lequel le système noradrénergique peut directement influencer les cibles corticales et être modulé par les projections glutamatergiques (Bouret et Sara, 2005 ; Briand et al., 2007 ; Sara, 2009). Par exemple, une étude récente suggère qu’il existe un « système d’alarme » reliant le tronc cérébral, l’amygdale et le cortex avec des connexions fonctionnelles entre le LC et l’ACC (Liddell et al., 2005).
Ainsi, s’il apparaît que la sérotonine puisse être liée à l’impulsivité et la noradrénaline à une augmentation de choix rapides qui ne tiennent pas compte des conséquences négatives à long terme, il ne faut pas oublier que ces deux neurotransmetteurs ne représentent qu’une petite partie des neurotransmetteurs, et qu’une explication ne tenant compte que d’un neurotransmetteur a de fortes chances d’être inexacte et incomplète.
Intégration et perspectives
La littérature sur l’imagerie cérébrale du suicide est peu développée. Les différentes études ont impliqué diverses parties du cortex préfrontal (la partie ventrolatérale (e.g., Leyton et al., 2006), incluant le cortex orbitofrontal (e.g., Monkul et al., 2007), la partie dorsomédiale (e.g., Oquendo et al., 2003) et la partie dorsolatérale (e.g., Audenaert et al., 2002)), l’ACC, l’amygdale et l’hippocampe (e.g., Monkul et al., 2007) (pour une revue voir Jollant et al., 2011)). Ces régions ont été impliquées dans une modulation défectueuse de l’attribution de valeurs et dans une régulation des émotions inefficace (Mikolajzak et Desseilles, 2012) pouvant conduire à faciliter un passage à l’acte dans certains contextes (Mikolajczak et Desseilles, 2012).
L’implication de régions cérébrales et de neurotransmetteurs, à la fois dans le processus suicidaire et dans la prise de décision (en particulier dans l’incertitude imprévue) peut suggérer un pattern de régions potentiellement impliquées dans le processus suicidaire qui devrait faire l’objet de recherches plus focalisées.
Récemment, des modèles complexes ont tenté d’intégrer les quatre principaux neuromodulateurs ascendants et plusieurs régions cérébrales dans l’évaluation et dans la décision (Doya, 2002 ; Doya, 2008). La dopamine signale l’erreur dans la prédiction de la récompense. Elle représente le signal global d’apprentissage pour la prédiction d’une récompense et le renforcement de l’action. La sérotonine contrôle l’échelle de temps de la prédiction de la récompense. Elle contrôle la balance entre la prédiction de la récompense à court terme ou à long terme. La noradrénaline contrôle l’aspect imprévisible de la sélection de l’action. Elle contrôle la balance entre une exploration large et une exécution focalisée. L’acétylcholine contrôle la vitesse de la mise à jour de la mémoire. Elle contrôle la balance entre le stockage en mémoire et le renouvellement.
Tableau 1
Quatre principaux neuromodulateurs ascendants, leurs origines et leurs principales cibles
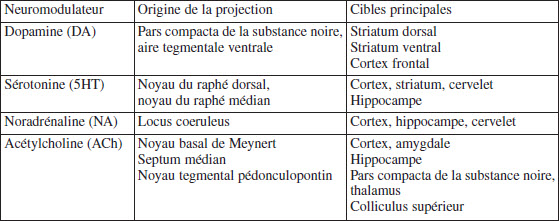
La vision parcellaire et les résultats parfois divergents des études visant à objectiver des marqueurs biologiques du processus suicidaire pourraient être mieux compris en intégrant les observations psychologiques et comportementales chez les patients suicidaires. Les théories d’évaluation et de décision (e.g. telles que proposées par Doya (2002)) pourraient également aider à la compréhension. Ces théories offrent de nombreux insights quant aux interactions entre les régions cérébrales potentiellement impliquées dans le processus suicidaire. En particulier, les quatre neuromodulateurs ascendants, l’origine de leurs projections et leurs cibles cérébrales principales (tableau 1) sont autant d’acteurs importants pour le processus suicidaire en tant qu’incertitude imprévue. En particulier, les études focalisant sur les noyaux des principaux neuromodulateurs et étudiant leur connectivité en rapport avec le processus suicidaire (idéation, plan, tentative) devraient permettre d’améliorer nos connaissances quant aux mécanismes cérébraux en cours dans le processus suicidaire, et ainsi ouvrir des pistes d’intervention et de prévention.
Nous proposons que l’étude des processus suicidaires au moyen de la métaphore de l’apprentissage par renforcement puisse permettre de mieux comprendre la physiopathologie du processus suicidaire et d’intégrer les résultats de la littérature existants.
Figure 3
Diagramme schématique et fonctions hypothétiques de l’apprentissage par renforcement, ses corrélats cérébraux et l’implication dans le processus suicidaire

Se référer au texte pour les détails.
Le but de l’apprentissage par renforcement est de trouver une politique optimale pour maximiser les récompenses attendues. Une structure classique utilisée pour décrire les facteurs en jeu est celle de l’acteur-critique (Barto et al., 1983 ; Barto, 1995). Cette configuration propose des algorithmes basés sur des statistiques incrémentales. L’apprentissage par renforcement est utile dans les cas où l’agent (critique et acteur) ne connaît pas la valeur des choix d’action et les récompenses associées. L’agent tient à jour une représentation de la fonction de valeur de la politique qu’il utilise et une représentation de cette politique. Une mise à jour de la fonction de valeur induit donc une mise à jour locale de la politique, et la prise d’informations sur l’environnement entraîne des mises à jour de l’agent (critique et acteur). Faisant suite à la figure 2, la figure 3 propose un diagramme schématique de l’apprentissage par renforcement et ses corrélats à des neuromodulateurs, tels que proposés par Doya (2002)
α est le taux d’apprentissage pour contrôler à quelle vitesse la mémoire ancienne est mise à jour par la nouvelle expérience. L’erreur de différence temporelle (δ) ou l’erreur de prédiction est utilisée comme signal d’erreur pour apprendre la fonction de valeur. Les récompenses — renforcements attendus dans un futur lointain — ont moins de poids et sont représentés par un paramètre γ, celui-ci exprimant donc la balance entre la prédiction de la récompense à court terme et à long terme. Un paramètre β contrôle la stochastique de la politique. Si β = 0, la sélection de l’action est aléatoire, si β = infini la sélection de l’action est déterminée en vue d’une récompense maximale. Pour les détails théoriques, se référer à l’article de Doya (2002).
Ce diagramme suggère que dans le cadre de l’étude des corrélats cérébraux du processus suicidaire, l’étude d’un seul neurotransmetteur ou d’une seule région cérébrale ne peut amener qu’à une compréhension partielle d’un mécanisme beaucoup plus englobant, c’est-à-dire l’attribution de valeur, le choix d’action et l’apprentissage par renforcement.
Conclusion
Loin d’être capables de prédire le suicide, les méthodes scientifiques actuelles permettent d’avoir une idée du niveau de risque suicidaire, qui lui-même peut orienter la prise en charge clinique des patients (Rogers et al., 2002). Des marqueurs psychologiques (Wenzel et al., 2008), biologiques (Mann et al., 2006) et comportementaux (Nock et al., 2010) ont été proposés. En résumé ces marqueurs ne sont que de faible puissance et ne se sont pas avérés, à ce jour, d’une grande utilité clinique dans la prévention du suicide.
Dans cet article, nous formulons l’hypothèse que la prédiction du suicide pourrait être rendue compliquée parce qu’elle résulte de la prédiction d’un choix, lequel est lui-même imprévisible. D’un point de vue biologique, l’incertitude imprévue serait déterminée par la noradrénaline et ses interactions avec la partie antérieure du cortex cingulaire (ACC).
Nous proposons dans cet article un modèle du suicide selon l’apprentissage par renforcement. Dans ce modèle, nous intégrons d’une part les quatre principaux modulateurs ascendants (acétylcholine, noradrénaline, sérotonine, dopamine) avec leurs régions de projections et d’afférences respectives, et d’autre part, différentes observations d’imagerie cérébrales trouvées à ce jour dans le processus suicidaire. Ce modèle s’appuie sur la proposition de Doya (2002), selon laquelle la dopamine représente le signal global d’apprentissage pour la prédiction d’une récompense et le renforcement de l’action ; la sérotonine contrôle la balance entre la prédiction de la récompense à court terme ou à long-terme ; la noradrénaline contrôle l’aspect imprévisible de la sélection de l’action et l’acétylcholine contrôle la vitesse de la mise à jour de la mémoire.
Ce modèle offre des perspectives d’étude et d’approfondissement de la physiopathologie du processus suicidaire, notamment via l’exploration systématique de l’impact des différents neuromodulateurs. Ceux-ci ne sont pas considérés dans notre modèle comme effecteurs uniques, mais plutôt en tant que paramètres interagissant dans l’équation complexe qui est celle de la prise de décision, du choix d’action, et de l’apprentissage par renforcement pouvant mener à l’acte suicidaire.
Parties annexes
Remerciements
Le support pour ce travail a été fourni par le Fonds national de la recherche scientifique. Un support additionnel a été fourni par l’Université de Liège, le Fonds Leon Fredericq, Wallonie-Bruxelles International et la Fondation Jean et Madeleine Vachoux. Les sponsors n’avaient pas de rôle dans le design, la collecte, l’analyse et l’interprétation des données, ni dans l’écriture du rapport et la décision de soumettre l’article pour publication.
Bibliographie
- Aston-Jones, G., Cohen, J. D., 2005, An integrative theory of locus coeruleus-norepinephrine function : adaptive gain and optimal performance, Annual Review of Neuroscience, 28, 403-50.
- Aston-Jones, G., Rajkowski, J., et al., 2000, Locus coeruleus and regulation of behavioral flexibility and attention, Progress in Brain Research, 126, 165-82.
- Aston-Jones, G., Rajkowski, J., et al., 1996, Role of the locus coeruleus in emotional activation, Progress in Brain Research, 107, 379-402.
- Audenaert, K., Goethals, I., et al., 2002, SPECT neuropsychological activation procedure with the Verbal Fluency Test in attempted suicide patients, Nuclear Medicine Communication, 23, 9, 907-16.
- Barto, A. G., 1995, Reinforcement learning in M. A. Arbib, ed., The Handbook of Brain Theory and Neural Networks, Cambridge, MA, MIT Press, 804-809.
- Barto, A. G., Sutton, R. S., et al., 1983, Neuronlike adaptive elements that can solve difficult learning control problems, IEEE Transactions on Systems, Man, and Cybernetics, 13, 834-846.
- Bayer, H. M., Glimcher, P. W., 2005, Midbrain dopamine neurons encode a quantitative reward prediction error signal, Neuron, 47, 1, 129-41.
- Becker, E. S., Strohbach, D., et al., 1999, A specific attentional bias in suicide attempters, Journal of Nervous and Mental Disease, 187, 12, 730-5.
- Behrens, T. E., Woolrich, M. W., et al., 2007, Learning the value of information in an uncertain world, Nature Neuroscience, 10, 9, 1214-21.
- Bouret, S., Sara, S. J., 2005, Network reset : a simplified overarching theory of locus coeruleus noradrenaline function, Trends in Neuroscience, 28, 11, 574-82.
- Briand, L. A., Gritton, H., et al., 2007, Modulators in concert for cognition : modulator interactions in the prefrontal cortex, Progress inNeurobiology, 83, 2, 69-91.
- Brown, G. K., Beck, A. T., et al., 2000, Risk factors for suicide in psychiatric outpatients : a 20-year prospective study, Journal of Consulting and Clinical Psychology, 68, 3, 371-7.
- Busch, K. A., Fawcett, J., et al., 2003, Clinical correlates of inpatient suicide, Journal of Clinical of Psychiatry, 64, 1, 14-9.
- Caspi, A., Sugden, K., et al., 2003, Influence of life stress on depression : moderation by a polymorphism in the 5-HTT gene, Science, 301, 5631, 386-9.
- Chiles, J. A., Strosahl, K. D., 2005, Clinical Manual for Assessment and Treatment of Suicidal Patients, American Psychiatric Publishing, Arlington, VA.
- D’Zurilla, T. J., Goldfried, M. R., 1971, Problem solving and behavior modification, Journal of Abnormal Psychology, 78, 1, 107-26.
- Daw, N. D., O’Doherty, J. P., et al., 2006, Cortical substrates for exploratory decisions in humans, Nature, 441, 7095, 876-9.
- Dayan, P., Kakade, S., et al., 2000, Learning and selective attention, Nature Neuroscience, 3, Suppl : 1218-23.
- Desseilles, M., Perroud, N., et al., 2011, Is it valid to measure suicidal ideation by depression rating scales ?, Journal of Affective Disorder, 136, 3, 398-404.
- Dombrovski, A. Y., Siegle, G. J., et al., 2011, The temptation of suicide : striatal gray matter, discounting of delayed rewards, and suicide attempts in late-life depression, Psychology Medicine, 1-13.
- Doya, K., 2002, Metalearning and neuromodulation, Neural Networks, 15- 4-6, 495-506.
- Doya, K., 2008, Modulators of decision making, Nature Neuroscience, 11, 4, 410-6.
- Eckhoff, P., Wong-Lin, K. F., et al., 2009, Optimality and robustness of a biophysical decision-making model under norepinephrine modulation, Journal of Neuroscience, 29, 13, 4301-11.
- Ernst, M. O., Banks, M. S., 2002, Humans integrate visual and haptic information in a statistically optimal fashion, Nature, 415, 6870, 429-33.
- Goldstein, R. B., Black, D. W., et al., 1991, The prediction of suicide. Sensitivity, specificity, and predictive value of a multivariate model applied to suicide among 1906 patients with affective disorders, Archives of General Psychiatry, 48, 5, 418-22.
- Haruno, M., Kuroda, T., et al., 2004, A neural correlate of reward-based behavioral learning in caudate nucleus : a functional magnetic resonance imaging study of a stochastic decision task, Journal of Neuroscience, 24, 7, 1660-5.
- Jollant, F., Lawrence, N. L., et al., 2011, The suicidal mind and brain : a review of neuropsychological and neuroimaging studies, World Journal of Biological Psychiatry, 12, 5, 319-39.
- Jollant, F., Lawrence, N. S., et al., 2008, Orbitofrontal cortex response to angry faces in men with histories of suicide attempts, American Journal of Psychiatry, 165, 6, 740-8.
- Jollant, F., Lawrence, N. S., et al., 2010, Decreased activation of lateral orbitofrontal cortex during risky choices under uncertainty is associated with disadvantageous decision-making and suicidal behavior, Neuroimage.
- Kahneman, D., Tversky, A., 2000, Choices, Values and Frames, Cambridge University Press.
- Kessler, R. C., Borges, G., et al., 1999, Prevalence of and risk factors for lifetime suicide attempts in the National Comorbidity Survey, Archives of General of Psychiatry, 56, 7, 617-26.
- Kording, K. P., Wolpert, D., M., 2004, Bayesian integration in sensorimotor learning, Nature, 427, 6971, 244-7.
- Kunishio, K., Haber, S. N., 1994, Primate cingulostriatal projection : limbic striatal versus sensorimotor striatal input, The Journal of Comparative Neurology, 350, 3, 337-56.
- Leyton, M., Paquette, V., et al., 2006, alpha- [11C] Methyl-L-tryptophan trapping in the orbital and ventral medial prefrontal cortex of suicide attempters, European Neuropsychopharmacoly, 16, 3, 220-3.
- Liddell, B. J., Brown, K. J., et al., 2005, A direct brainstem-amygdala-cortical “alarm” system for subliminal signals of fear, Neuroimage, 24, 1, 235-43.
- Luksys, G., Gerstner, W., et al., 2009, Stress, genotype and norepinephrine in the prediction of mouse behavior using reinforcement learning, Nature Neuroscience, 12, 9, 1180-6.
- Mancini, C., Brown, G. M., 1992, Urinary catecholamines and cortisol in parasuicide, Psychiatry Research, 43, 1, 31-42.
- Mann, J. J., 2003, Neurobiology of suicidal behavior, Nature Reviews Neuroscience, 4, 10, 819-28.
- Mann, J. J., Currier, D., et al., 2006, Can biological tests assist prediction of suicide in mood disorders ? International Journal of Neuropsychopharmacoly, 9, 4, 465-74.
- Marchand, W. R., Lee, J. N., et al., 2012, Striatal and cortical midline circuits in major depression : implications for suicide and symptom expression, Progress in Neuropsychopharmacoly and Biological Psychiatry, 36, 2, 290-9.
- McCoy, A. N., Crowley, J. C., et al., 2003, Saccade reward signals in posterior cingulate cortex, Neuron, 40, 5, 1031-40.
- Mikolajczak, M., Desseilles, M., 2012, Traité de régulation des émotions, Bruxelles, de Boeck.
- Monkul, E. S., Hatch, J. P., et al., 2007, Fronto-limbic brain structures in suicidal and non-suicidal female patients with major depressive disorder, Molecular Psychiatry, 12, 4,360-6.
- Montague, P. R., Dayan, P., et al., 1995, Bee foraging in uncertain environments using predictive hebbian learning, Nature, 377, 6551, 725-8.
- Montague, P. R., King-Casas, B., et al., 2006, Imaging valuation models in human choice, Annual Review of Neuroscience, 29, 417-48.
- Nock, M. K., Banaji, M. R., 2007, Prediction of suicide ideation and attempts among adolescents using a brief performance-based test, Journal of Consulting and Clinical Psychology, 75, 5, 707-15.
- Nock, M. K., Borges, G., et al., 2008, Cross-national prevalence and risk factors for suicidal ideation, plans and attempts, British Journal of Psychiatry, 192, 2, 98-105.
- Nock, M. K., Park, J. M., et al., 2010, Measuring the suicidal mind : implicit cognition predicts suicidal behavior, Psychological Science, 21, 4, 511-7.
- O’Doherty, J., Dayan, P., et al., 2004, Dissociable roles of ventral and dorsal striatum in instrumental conditioning, Science, 304, 5669, 452-4.
- Oldershaw, A., Grima, E., et al., 2009, Decision making and problem solving in adolescents who deliberately self-harm, Psychological Medicine, 39, 1, 95-104.
- Oquendo, M. A., Galfalvy, H., et al., 2004, Prospective study of clinical predictors of suicidal acts after a major depressive episode in patients with major depressive disorder or bipolar disorder, American Journal of Psychiatry, 161, 8, 1433-41.
- Oquendo, M. A., Placidi, G. P., et al., 2003, Positron emission tomography of regional brain metabolic responses to a serotonergic challenge and lethality of suicide attempts in major depression, Archives of General Psychiatry, 60, 1, 14-22.
- Perroud, N., Aitchison, K. J., et al., 2009, Genetic Predictors of Increase in Suicidal Ideation During Antidepressant Treatment in the GENDEP Project, Neuropsychopharmacology.
- Qin, P., Nordentoft, M., 2005, Suicide risk in relation to psychiatric hospitalization : evidence based on longitudinal registers, Archives of General Psychiatry, 62, 4, 427-32.
- Rescorla, R. A., Wagner, A. R., 1972, A theory of Pavlovian conditioning : variations in the reinforcement and nonreinforcement, Classical Conditioning II : Current Research and Theory, A. H. Black, Prokasy, W. F., New York, Appleton-Century Crofts, 64-99.
- Risch, N., Herrell, R., et al., 2009, Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression : a meta-analysis, JAMA, 301, 23, 2462-71.
- Rogers, J. R., Lewis, M. M., et al., 2002, Validity of the suicide assessment checklist in an emergency crisis center, Journal of Counseling and Development, 80, 493-502.
- Samejima, K., Ueda, Y., et al., 2005, Representation of action-specific reward values in the striatum, Science, 310, 5752, 1337-40.
- Sara, S. J., 2009, The locus coeruleus and noradrenergic modulation of cognition, Nature Reviews Neuroscience, 10, 3, 211-23.
- Shneidman, E., 1985, Definition of Suicide, John Wiley and Sons, New York.
- Sutton, R. S., Barto, A. G., 1998, Reinforcement Learning : an Introduction, Cambridge, Massachusetts, MIT Press.
- TANAKA, S. C., DOYA, K., et al., 2004, Prediction of immediate and future rewards differentially recruits cortico-basal ganglia loops, Nature Neuroscience, 7, 8, 887-93.
- Waelti, P., Dickinson, A., et al., 2001, Dopamine responses comply with basic assumptions of formal learning theory, Nature, 412, 6842, 43-8.
- Wenzel, A., Brown, Et Al., G. K., 2008, Cognitive Therapy for Suicidal Patients : Scientific and Clinical Applications, American Psychological Association (APA), Washington.
- Wenzel, A., Brown, G. K., et al., 2009, Cognitive Therapy for Suicidal Patients : Scientific and Clinical Applications, American Psychological Association (APA), Washington.
- West, C. H., Ritchie, J. C., et al., 2010, Paroxetine-induced increase in activity of locus coeruleus neurons in adolescent rats : implication of a countertherapeutic effect of an antidepressant, Neuropsychopharmacology, 35, 8, 1653-63.
- Williams, J. M., Broadbent, K., 1986, Distraction by emotional stimuli : use of a Stroop task with suicide attempters, British Journal of Clinical Psychology, 25, 101-10.
- Williams, J. M. G., Barnhoffer, T., et al., 2006, The role of overgeneral memory in suicidality, Cognition and Suicide : Theory, Research, and Therapy, T. E. Ellis. Washington, DC, American Psychiatric Association : 173-192.
- Yu, A. J., Dayan, P., 2005, Uncertainty, neuromodulation, and attention, Neuron, 46, 4, 681-92.
Liste des figures
Figure 1
Déficits pour la résolution de problèmes
Les stress de notre vie et les tracas quotidiens agissent sur nos processus cognitifs et en particulier sur les cognitions en rapport avec le suicide, sous forme de pensées ou d’interprétations. Ces modifications cognitives influent sur le traitement de l’information, premièrement à court terme, via des biais attentionnels qui amènent le sujet à diriger son attention vers des stimuli négatifs, deuxièmement à long terme, via des biais mnésiques résultant d’une mémoire surgénéralisée. En particulier, la fixation attentionnelle et la constriction cognitive entraînent des déficits pour la résolution de problèmes.
Figure 2
Théorie de l’apprentissage
Selon le modèle d’apprentissage par renforcement (Sutton et Barto, 1998), lorsque nous recevons de nouvelles informations, nous mettons à jour nos croyances à propos de l’environnement, en proportion de l’erreur de prédiction (c’est-à-dire la différence entre l’issue escomptée et l’issue actuelle d’une action, le δ sur la figure). Ce doit être multiplié par un facteur (α) appelé le taux d’apprentissage, afin de déterminer le degré par lequel la valeur de l’action est mise à jour.
Figure 3
Diagramme schématique et fonctions hypothétiques de l’apprentissage par renforcement, ses corrélats cérébraux et l’implication dans le processus suicidaire
Se référer au texte pour les détails.
Liste des tableaux
Tableau 1
Quatre principaux neuromodulateurs ascendants, leurs origines et leurs principales cibles