Abstracts
Résumé
La coccinelle asiatique, Harmonia axyridis, une espèce introduite, établie au Québec depuis 1994, est rapidement devenue un problème entomologique puisqu’elle s’abrite dans les bâtiments durant la période hivernale. L’une des raisons souvent invoquées pour expliquer ce comportement serait une faible résistance au froid de cette espèce. Pour déterminer la résistance au froid des adultes de la coccinelle asiatique, une expérience combinant à la fois des températures froides et différentes durées d’exposition a été réalisée. Des groupes de 50 coccinelles placées dans des plats de Pétri ont été exposés à différentes températures (5, 0, -5, -10 et –20oC) pour des périodes de temps prédéterminées (2, 6, 12 et 18 semaines). Les résultats indiquent que le taux de survie est à la fois influencé par la température et la durée d’exposition. De plus, aucune coccinelle asiatique n’a survécu à –20oC, même après une exposition d’une seule journée.
Abstract
The Asian ladybird beetle, Harmoniaaxyridis, an introduced species that is established in Quebec since 1994, rapidly became an entomological problem because it aggregates in buildings during fall and winter. The reason most often invoked to explain this behaviour is a weak resistance of the species to winter coldness. To determine cold resistance of the Asian ladybird beetle, a study including various cold temperatures and exposure times was carried out. Groups of 50 beetles were placed in Petri dishes and exposed to different temperatures (5, 0, -5, -10 and –20oC) for different periods of time (2, 6, 12 and 18 weeks). Results indicate that survival is strongly influenced by temperature but also by exposure duration. Furthermore, no Asian ladybird beetle survived at –20oC, even for only one day of exposure.
Article body
Introduction
La coccinelle asiatique, Harmoniaaxyridis (Pallas) [Coleoptera : Coccinellidae], est un prédateur généraliste qui a été introduit comme agent de lutte biologique à plusieurs reprises aux États-Unis durant le siècle dernier avec plus ou moins de succès (McClure 1986; Tedders et Schaefer 1994). La majorité de ces introductions n’auraient pas permis l’établissement de cette coccinelle sur le continent nord-américain. Finalement, Chapin et Brou (1991) ont décrit pour la première fois, l’intégration de la coccinelle asiatique à l’entomofaune d’Amérique du Nord en Louisiane, en 1988. Cette espèce exotique a rapidement envahi l’ensemble du continent nord-américain (Tedders et Schaefer 1994) en comparaison avec d’autres espèces de coccinelles exotiques introduites précédemment (McCorquodale 1998). Au Québec, la première mention de cette coccinelle date de 1994 (Coderre et al. 1995), soit seulement 6 ans après son établissement aux États-Unis.
La coccinelle asiatique est considérée comme un insecte bénéfique et un prédateur hautement efficace (Dixon 2000), bien qu’aucune étude n’ait véritablement démontré un impact majeur sur les ravageurs (Boiteau et al. 1999; Wallace et Hain 2000). Par ailleurs, en Amérique du Nord, cette coccinelle est désormais considérée par plusieurs comme un insecte nuisible puisqu’elle se réfugie par dizaines de milliers à l’intérieur des habitations durant la saison hivernale (Nalepa et al. 1996, 2000; Riddick et al. 2000; Tedders et Schaefer 1994) et peut même provoquer des réactions allergiques (rhino-conjonctivites) chez les occupants (Yarbrough et al. 1999).
Les raisons expliquant ce phénomène d’agrégation dans les bâtiments lors de l’approche de la saison hivernale demeurent encore aujourd’hui nébuleuses. Cependant, l’une des raisons invoquées pour expliquer ce phénomène serait une faible résistance de l’espèce aux froides températures hivernales (Berthiaume 1999; Giroux 2002) puisque les premiers essais effectués indiquaient que le taux de mortalité d’adultes placés dans des cages sous la surface du sol au Connecticut dépassait 95 % (McClure 1987). La présente étude a pour objectif d’évaluer expérimentalement l’impact de la température et de la durée d’exposition au froid sur les capacités de survie de cette espèce.
Matériel et méthodes
Les 11 et 12 octobre 2001, plus de 5000 coccinelles asiatiques ont été capturées à la main sur un bâtiment de la région de Québec (46o46’N; 71o17’O) lors du déplacement des coccinelles vers leur site potentiel d’hibernation. Tous les spécimens capturés ont été gardés au froid (4oC) durant quelques jours dans des contenants de plastique de 300 mL, par groupes d’environ 100 individus par contenant. Cette courte période d’exposition à une basse température a permis de manipuler et de dénombrer les coccinelles afin de les distribuer par groupe de 50 individus à l’intérieur de plats de Pétri de 145 mm de diam. Par la suite, les plats de Pétri ont été entreposés dans une chambre de croissance à une température de 8oC et une photopériode de 10L:14O, réduisant ainsi fortement l’activité des coccinelles afin de minimiser les pertes énergétiques avant le début des traitements au froid. La température de la chambre de croissance a été progressivement diminuée d’un degré par semaine jusqu’à 4oC durant le mois suivant.
Au début de décembre, les plats de Pétri contenant les coccinelles ont été transférés dans cinq chambres froides ayant chacune des conditions thermiques différentes et constantes. À l’intérieur des chambres froides, les coccinelles étaient placées à l’obscurité complète afin de reproduire les conditions d’hibernation à l’intérieur des maisons (grenier, espace entre les murs, etc.). Les températures testées durant l’expérimentation ont été les suivantes : -20, -10, -5, 0 et 5 ± 1oC. L’humidité relative a été maintenue constante à environ 30 % à l’intérieur des chambres froides. Dans chaque chambre froide, 16 plats de Pétri contenant chacun 50 coccinelles ont été disposés de manière aléatoire. Afin d’évaluer l’effet de la température et de la durée de l’exposition au froid sur la survie des coccinelles, quatre plats de Pétri ont été retirés de manière aléatoire après 2, 6, 12 et 18 semaines consécutives d’exposition à la même température. Après chaque durée d’exposition, les plats de Pétri ont été retirés des chambres froides et placés à la température de la pièce (22 ± 1oC). Durant les 5 j suivant la sortie des chambres froides, les coccinelles capables de se déplacer ont été retirées, dénombrées et considérées comme viables, alors que celles qui n’ont montré aucun signe d’activité après 5 j ont été considérées comme ayant péri au cours du traitement.
Une autre expérience a été réalisée pour démontrer l’effet de courtes périodes d’exposition (1, 2, 4 et 8 j) à –20°C sur la survie de la coccinelle asiatique. Durant cette expérience, les facteurs abiotiques ont été maintenus constants pour chaque période d’exposition. Quatre plats de Pétri contenant 40 coccinelles chacun ont été exposés pour chaque période d’exposition testée, puis exposés à la température de la pièce après avoir été retirés de la chambre froide. Les mêmes critères énoncés précédemment ont été utilisés afin d’identifier les individus morts et vivants.
La survie de la coccinelle asiatique a également été évaluée à une température constante de 5oC pour une durée d’exposition de 40 semaines consécutives afin d’évaluer les capacités de cette coccinelle dans des conditions extrêmes comme celles qu’elle rencontrerait dans les habitations du nord du Québec, où l’hiver est plus long. Pour cette expérience, six plats de Pétri contenant chacun 50 coccinelles ont été placés à l’obscurité pendant 40 semaines dans une chambre de croissance à 5oC. Après cette période, les plats de Pétri ont été retirés et exposés à la température de la pièce. Les mêmes critères énoncés précédemment ont été utilisés afin d’identifier les individus morts et vivants.
Analyses statistiques
Une analyse de variance à deux critères de classification a été utilisée pour déterminer l’effet de la température et de la durée d’exposition au froid ainsi que l’interaction possible de ces deux facteurs sur le taux de survie de la coccinelle asiatique. Les pourcentages de survie ont été transformés en utilisant la fonction arcsinš%survie pour respecter les postulats de l’analyse de variance (homogénéité des variances). Une analyse de variance a également été utilisée pour chaque température testée. Cette dernière était suivie d’un test de comparaisons multiples LSD de Fisher lorsque l’analyse de variance était significative afin de déterminer quelles durées d’exposition montraient des différences significatives pour une température donnée. Les analyses ont été effectuées avec le logiciel SAS (1999).
Résultats
Les résultats indiquent une interaction hautement significative (F = 9,59; d.l. = 12, 60; P < 0,0001) entre la température et la durée d’exposition sur le taux de survie des adultes de la coccinelle asiatique (Tableau 1), ce qui implique que l’effet de la durée d’exposition n’est pas toujours le même selon la température considérée. Selon le modèle testé, plus de 93 % de la variance est expliquée par les deux facteurs considérés soient la température et la durée d’exposition.
Tableau 1
Résultat de l’analyse de variance sur l’effet de la température et de la durée d’exposition sur le taux de survie de la coccinelle asiatique Harmoniaaxyridis

Pour les expositions à 5 et 0oC (Figs. 1a et 1b), aucune différence significative (5oC : F = 1,96; d.l. = 3, 12; P = 0,1741; 0oC : F = 0,14; d.l. = 3, 12; P = 0,9340) n’a été observée entre les différentes durées d’exposition. La survie des coccinelles asiatiques exposées à ces deux températures est toujours supérieure à 80 % peu importe la durée d’exposition. Les résultats obtenus pour une période d’exposition de 40 semaines à 5oC indiquent, cependant, que seulement 22,3 ± 1,9 % (moyenne ± erreur type) des coccinelles survivent à un traitement aussi prolongé.
Pour l’exposition à –5oC (Fig. 1c), une différence significative du taux de survie des coccinelles (F = 10,15; d.l. = 3, 12; P = 0,0013) est observée en fonction de la durée d’exposition. Aucune différence n’a cependant été observée dans la survie de la coccinelle asiatique aux trois premières périodes d’exposition (2, 6 et 12 semaines), la survie demeurant toujours supérieure à 80 % comme pour les températures de 5 et 0oC. Cependant, seulement 66,5 % des coccinelles ont survécu à une exposition de 18 semaines à –5oC (Fig. 1c).
Pour une température de –10oC, le taux de survie de la coccinelle asiatique atteint 88 % après 2 semaines d’exposition mais diminue rapidement à 13 % après 6 semaines d’exposition (Fig. 1d) (F = 118,14; d.l. = 3, 12; P < 0,0001). Aucune coccinelle n’a survécu à une exposition de 12 ou 18 semaines à cette température. Par ailleurs, les résultats indiquent clairement que H. axyridis est incapable de survivre à une température de –20oC puisque aucune coccinelle n’a été retrouvée vivante pour chacune des quatre durées d’exposition testées. Les résultats des courtes périodes d’exposition (1, 2, 4 et 8 j) à cette même température (–20oC) ont également démontré que toutes les coccinelles périssent lorsqu’elles y sont exposées.
Figure 1
Taux de survie de la coccinelle Harmoniaaxyridis à différentes températures et pour différentes périodes d’exposition
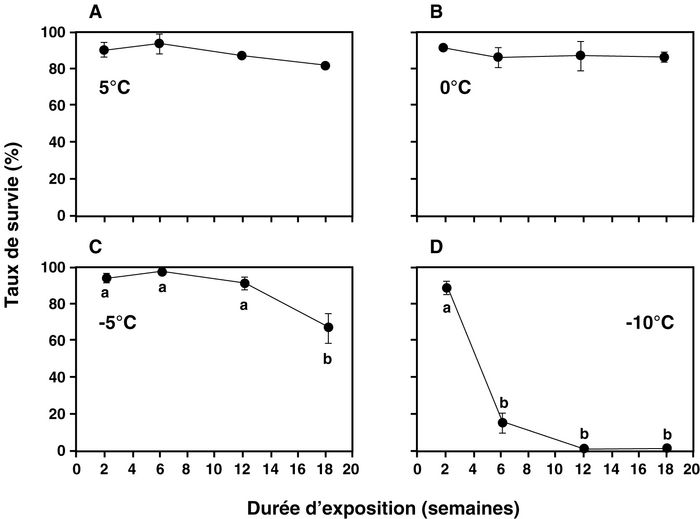
Des lettres différentes indiquent des différences significatives de survie entre les périodes d’exposition pour une même température.
Discussion
La survie hivernale d’une espèce dépend non seulement de sa capacité à résister aux diminutions de la température ambiante mais également des stratégies qu’elle déploie pour atténuer les impacts négatifs de cette diminution de température (Leather et al. 1993; Lee et Denlinger 1991). Les stratégies déployées par les espèces peuvent être comportementales ou physiologiques (Leather et al. 1993; Lee et Denlinger 1991). Chez la coccinelle asiatique, l’une des stratégies comportementales utilisées est d’effectuer des agrégations dans des endroits où l’humidité relative est peu importante comme les parois rocheuses (Obata 1986; Sakurai et al. 1993) et les bâtiments. Ce comportement d’agrégation à l’intérieur des bâtiments est maintenant largement répandu en Amérique du Nord (Nalepa et al. 2000; Riddick et al. 2000; Tedders et Schaefer 1994) mais n’est pas exclusif à ce continent puisque la coccinelle asiatique effectue également ce type d’agrégation dans son aire de distribution d’origine (Sakurai et al. 1993; Watanabe 2002).
La température a une influence majeure sur la survie hivernale des insectes mais il faut également considérer la durée d’exposition au froid lorsqu’on évalue la survie hivernale d’une espèce (Jean et al. 1990; Salt 1961). Nos résultats indiquent clairement que la température influence le taux de survie de la coccinelle asiatique mais que la durée d’exposition joue également un rôle important. De plus, l’effet de la durée d’exposition diffère selon les températures testées. En effet, aucun effet de la durée d’exposition n’a été observé pour les deux températures les plus chaudes (5 et 0oC) alors que ce facteur a eu un effet important lorsque les individus ont été exposés à des températures de –5 et –10oC. L’effet de la température dépend donc inévitablement de la durée d’exposition (Lee et Denlinger 1991; Salt 1961) puisque même à 5oC, on observe une diminution de la survie des coccinelles lorsqu’elles sont exposées à cette température pour une très longue période de temps (40 semaines).
Les taux de survie des coccinelles asiatiques lorsqu’elles ont été exposées à des températures de 0 et –5oC en conditions expérimentales sont comparables à ceux obtenus par Watanabe (2002) au Japon, soit environ 90 %, pour des durées d’exposition similaires. Par ailleurs, les individus testés durant notre expérience ont montré moins de résistance au froid que ceux étudiés par Voronine (1965) en Russie qui rapporte 100 % de survie chez des individus exposés à une température de –10oC pendant 2 mois. Nos résultats montrent une survie de seulement 13 % pour des individus exposés à –10oC pendant 6 semaines. Cet important écart entre ces deux études peut être le résultat de différences entre les deux populations de coccinelles testées. En effet, la population testée par Voronine (1965) provenait de la Sibérie, un milieu beaucoup plus froid que celui d’où originerait la souche nord-américaine de la coccinelle asiatique. En effet, la plupart des relâchers de la coccinelle asiatique ont été réalisés avec des individus qui provenaient du Japon qui possède un climat plus tempéré que celui de la Sibérie. D’ailleurs, la coccinelle asiatique montre une grande variabilité génétique (Majerus et al. 1998; McClure 1987) qui pourrait dissimuler une variation régionale dans ses capacités de résistance au froid.
La résistance au froid de la coccinelle asiatique est supérieure à celle de la coccinelle maculée (Coleomegillamaculatalengi Timberlake), une espèce indigène à l’Amérique du Nord. En effet, Jean et al. (1990) ont réalisé des expériences avec la coccinelle maculée à des températures et durées d’exposition semblables à celles de la présente étude sauf que les coccinelles étaient conservées dans des contenants avec un substrat imitant la litière du sol. Dans la présente étude, les coccinelles asiatiques étaient directement exposées aux températures et ne bénéficiaient donc d’aucune protection thermique. En réalité, des différences importantes existent entre ces deux espèces lorsque les températures extrêmes sont comparées pour des périodes d’exposition similaires. Environ 85 % des coccinelles asiatiques ont survécu à 18 semaines d’exposition à 5oC, comparativement à seulement 15 % pour les coccinelles maculées (Jean et al. 1990). Cette différence du taux de survie est également évidente lorsque les deux espèces sont exposées à une température de -10oC. Ainsi, près de 90 % des coccinelles asiatiques ont survécu à 2 semaines d’exposition à cette température comparativement à seulement 18 % chez la coccinelle maculée (Jean et al. 1990). Pour les températures intermédiaires (0 et –5 oC), le taux de survie est comparable entre les deux espèces bien que la coccinelle asiatique ne bénéficiait d’aucune protection thermique. Ainsi, les résultats indiquent que la capacité de survie au froid de la coccinelle asiatique en milieu expérimental est supérieure à celle de la coccinelle maculée bien que cette dernière soit une espèce indigène et donc bien adaptée aux conditions climatiques de l’Amérique du Nord.
La résistance élevée au froid de la coccinelle asiatique pourrait lui conférer un avantage sur les coccinelles indigènes si un réchauffement climatique entraînant une réduction de la couverture nivale se concrétisait. En effet, les espèces passant l’hiver au sol seraient alors exposées à des températures beaucoup plus variables que la coccinelle asiatique qui passe l’hiver à l’intérieur des murs et des greniers des habitations ainsi que dans les parois rocheuses en milieu naturel. La coccinelle asiatique possède aussi plusieurs autres caractéristiques (fécondité élevée, polymorphisme, polyphagie, etc.) qui pourraient l’avantager dans la compétition avec les autres espèces de coccinelles indigènes (Hodek et Honek 1996; Nalepa et al. 1996; Tedders et Schaefer 1994), ce qui pourrait expliquer son succès dans la colonisation de l’Amérique et représenter un risque pour les espèces de coccinelles indigènes.
Les faibles taux de survie hivernale rapportés par McClure (1987) lors des premiers essais réalisés en Amérique du Nord pourraient être le résultat de l’approche expérimentale utilisée. En effet, dans son expérience, McClure (1987) a placé les coccinelles dans la litière du sol, un environnement humide qui ne correspondrait pas aux sites d’hibernation naturels de la coccinelle asiatique, contrairement à plusieurs de nos espèces indigènes (Hodek et Honek 1996). En effet, la coccinelle asiatique préfère les parois rocheuses (Obata 1986; Sakurai et al. 1993), où l’humidité relative est beaucoup moins élevée que dans la litière du sol. L’hypothèse d’une faible résistance au froid de la coccinelle asiatique, souvent invoquée pour expliquer le phénomène d’agrégation à l’intérieur des bâtiments (Berthiaume 1999; Giroux 2002), n’est donc pas supportée par nos résultats, qui, comme ceux de Voronine (1965), indiquent que cette espèce possède les attributs nécessaires pour résister au froid.
Appendices
Remerciements
La réalisation de cette étude a été rendue possible grâce à la collaboration de Luc Lamontagne, Sylvain Lamontagne et Olivier Desbiens du Centre de foresterie des Laurentides du Service canadien des forêts et de Martin Charest du laboratoire d’entomologie forestière de l’Université Laval.
Références
- Berthiaume, R. 1999. La face cachée de la coccinelle exotique Harmonia axyridis. Antennae 6 : 5-9.
- Boiteau, G., Y. Bousquet et W.P.L. Osborn. 1999. Vertical and temporal distribution of Coccinellidae (Coleoptera) in flight over an agricultural landscape. Can. Entomol. 131 : 269-277.
- Chapin, J.B. et V.A. Brou. 1991. Harmoniaaxyridis (Pallas), the third species of the genus to be found in the United States (Coleoptera : Coccinellidae). Proc. Entomol. Soc. Wash. 93 : 630-635.
- Coderre, D., É. Lucas et I. Gagné. 1995. The occurence of Harmonia axyridis (Pallas) (Coleoptera : Coccinellidae) in Canada. Can. Entomol. 127 : 609-611.
- Dixon, A.F.G. 2000. Insect predator-prey dynamics: Ladybird beetles and biological control. Cambridge University Press, Cambridge. 257 pp.
- Giroux, M. 2002. Insectes utiles ou nuisibles? Bulletin de l’Entomofaune 26 : 9-11.
- Hodek, I. et A. Honek. 1996. Ecology of Coccinellidae. Kluwer Academic Publishers, Boston. 464 pp.
- Jean, C., D. Coderre et J.C. Tourneur. 1990. Effects of temperature and substrate on survival and lipid consumption of hibernating Coleomegillamaculatalengi (Coleoptera : Coccinellidae). Environ. Entomol. 19 : 1657-1662.
- Leather, S.R., K.F.A. Walters et J.S. Bale. 1993. The ecology of insect overwintering. Cambridge University Press, Cambridge. 255 pp.
- Lee, R.E. et D.L. Denlinger. 1991. Insects at low temperature. Chapman and Hall, NY. 508 pp.
- Majerus, T.M.O., M.E.N. Majerus, B. Knowles, J. Wheeler, D. Bertrand, V.N. Kuznetzov, H. Ueno et G.D.D. Hurst. 1998. Extreme variation in the prevalence of inherited male-killing microorganisms between three populations of Harmoniaaxyridis (Coleoptera : Coccinellidae). Heredity 81 : 683-691.
- McClure, M.S. 1986. Importing ladybird beetles to control red pine scale. Front. Plant Sci. 39 : 5-7.
- McClure, M.S. 1987. Potential of the asian predator Harmoniaaxyridis Pallas (Coleoptera : Coccinellidae), to control Matsucoccusresinosae Bean and Godwin (Homoptera : Margarodidae) in the United States. Environ. Entomol. 16 : 224-230.
- McCorquodale, D.B. 1998. Adventive lady beetles (Coleoptera : Coccinellidae) in eastern Nova Scotia, Canada. Entomol. News 109 : 15-20.
- Nalepa, C.A., K.A. Kidd et K.R. Ahlstrom. 1996. Biology of Harmoniaaxyridis (Coleoptera : Coccinellidae) in winter aggregations. Ann. Entomol. Soc. Am. 89 : 681-685.
- Nalepa, C.A., K.A. Kidd et D.I. Hopkins. 2000. The multicolore asian lady beetle (Coleoptera : Coccinellidae): Orientation to aggregation sites. J. Entomol. Sci. 35 : 150-157.
- Obata, S. 1986. Determination of hibernation site in the ladybird beetle, Harmoniaaxyridis Pallas (Coleoptera : Coccinellidae). Kontyû 54 : 218-223.
- Riddick, E.W., J.R. Aldrich, A.D. Milo et J.C. Davis. 2000. Potential for modifying the behavior of the multicoloured asian lady beetle (Coleoptera : Coccinellidae) with plant-derived natural products. Ann. Entomol. Soc. Am. 93 : 1314-1321.
- Sakurai, H., Y. Kumada et S. Takeda. 1993. Seasonal prevalence and hibernating- diapause behavior in the lady beetle, Harmoniaaxyridis. Res. Bull. Fac. Agric. Gifu Univ. 58 : 51-55.
- Salt, R.W. 1961. Principals of insect cold-hardiness. Annu. Rev. Entomol. 6 : 55-74.
- SAS Institute Inc. 1999. SAS/STAT® User’s Guide. Version 8. Volumes 1, 2 et 3. Cary. NC. 3884 pp.
- Tedders, W.L. et P.W. Schaefer. 1994. Release and establishment of Harmoniaaxyridis (Coleoptera : Coccinellidae) in the southeastern United States. Entomol. News 105 : 228-243.
- Voronine, E.K. 1965. Particularités de la formation de la population hibernale chez Harmoniaaxyridis, prédateur des aphidiens en Extrême-Orient. Trav. Inst. Prot. Plant. 24 : 227-234.
- Wallace, M.S. et F.P. Hain. 2000. Field surveys and evaluation of native and established predators of the hemlock wooly adelgid (Homoptera : Adelgidae) in the southeastern United States. Environ. Entomol. 29 : 638-644.
- Watanabe, M. 2002. Cold tolerance and myo-inosytol accumulation in overwintering adults of a lady beetle, Harmoniaaxyridis (Coleoptera : Coccinellidae). Eur. J. Entomol. 99 : 5-9.
- Yarbrough, J.A., J.L. Armstrong, M.Z. Blumberg, A.E. Phillips, E. McGahee et W.K. Dolen. 1999. Allergic rhinoconjunctivitis caused by Harmonia axyridis (Asian lady beetle, Japanese lady beetle or lady bug). J. Allergy Clin. Immunol. 104 : 704-705.
List of figures
Figure 1
Taux de survie de la coccinelle Harmoniaaxyridis à différentes températures et pour différentes périodes d’exposition
Des lettres différentes indiquent des différences significatives de survie entre les périodes d’exposition pour une même température.
List of tables
Tableau 1
Résultat de l’analyse de variance sur l’effet de la température et de la durée d’exposition sur le taux de survie de la coccinelle asiatique Harmoniaaxyridis